

WLS/wntless is essential in controlling dendritic cell homeostasis via a WNT signaling-independent mechanism
- PMID: 33853474
- PMCID: PMC8726611
- DOI: 10.1080/15548627.2021.1907516
WLS/wntless is essential in controlling dendritic cell homeostasis via a WNT signaling-independent mechanism
Abstract
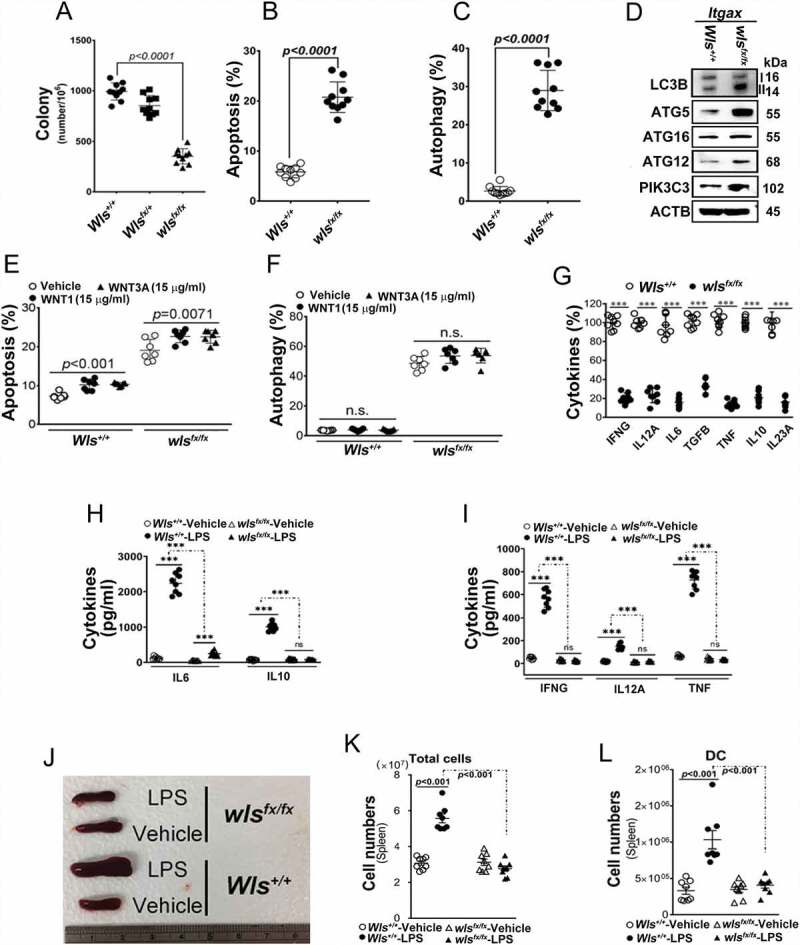
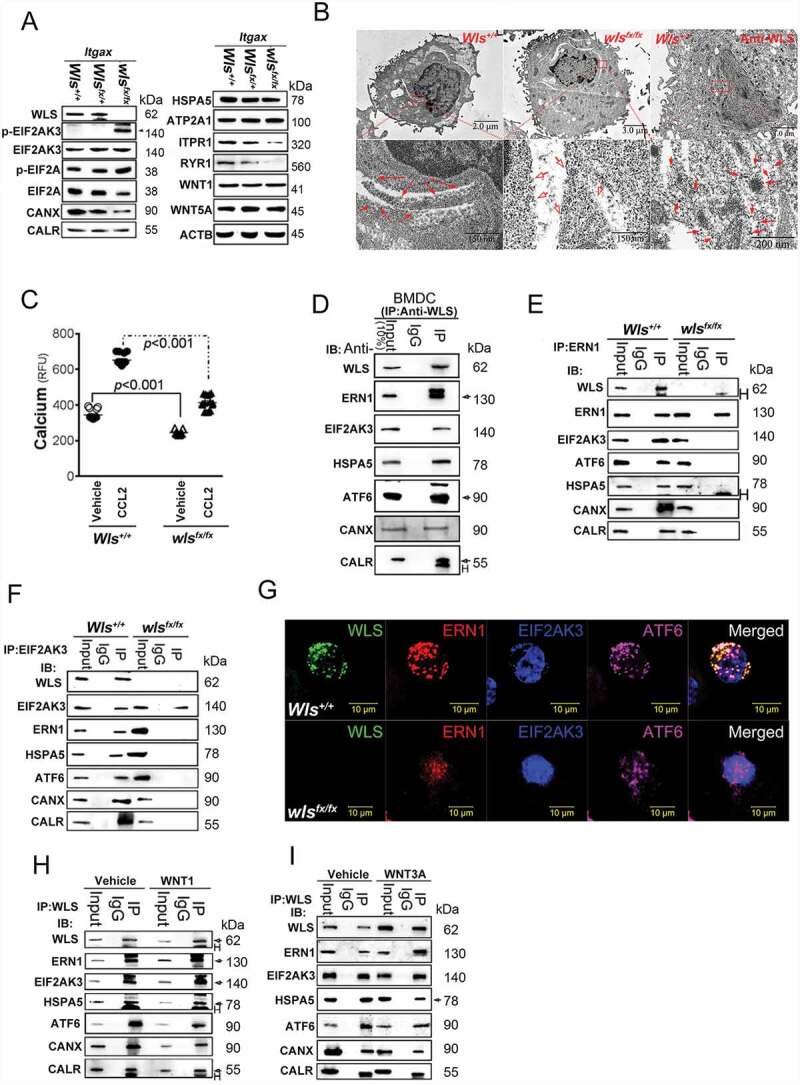
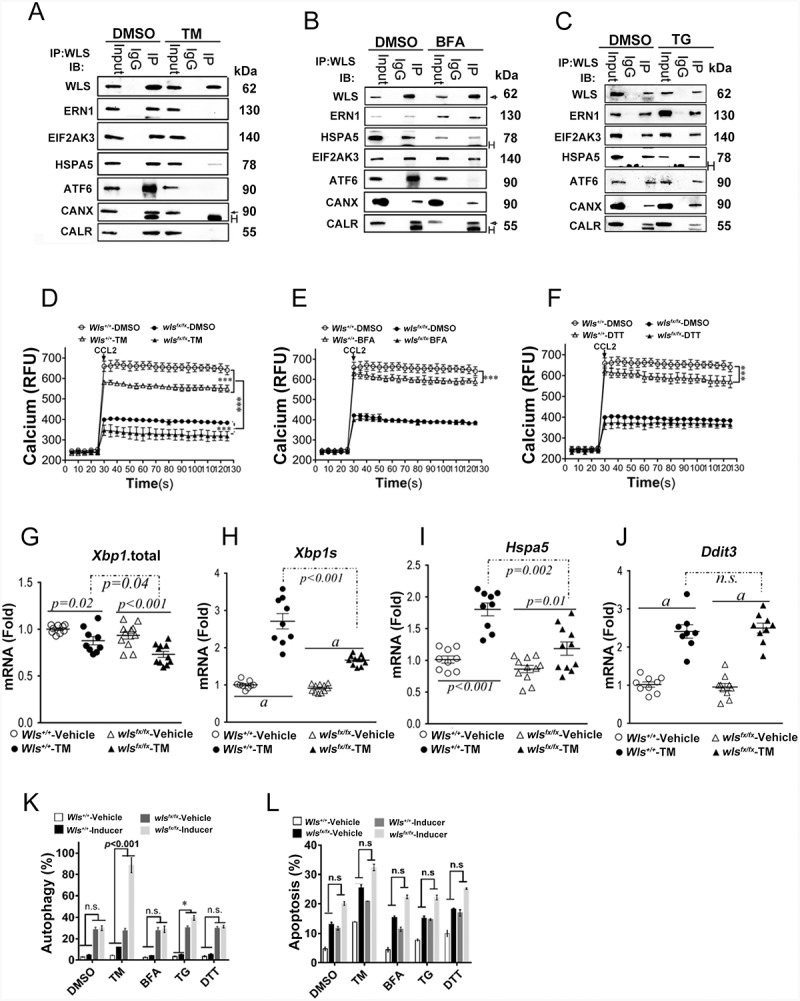
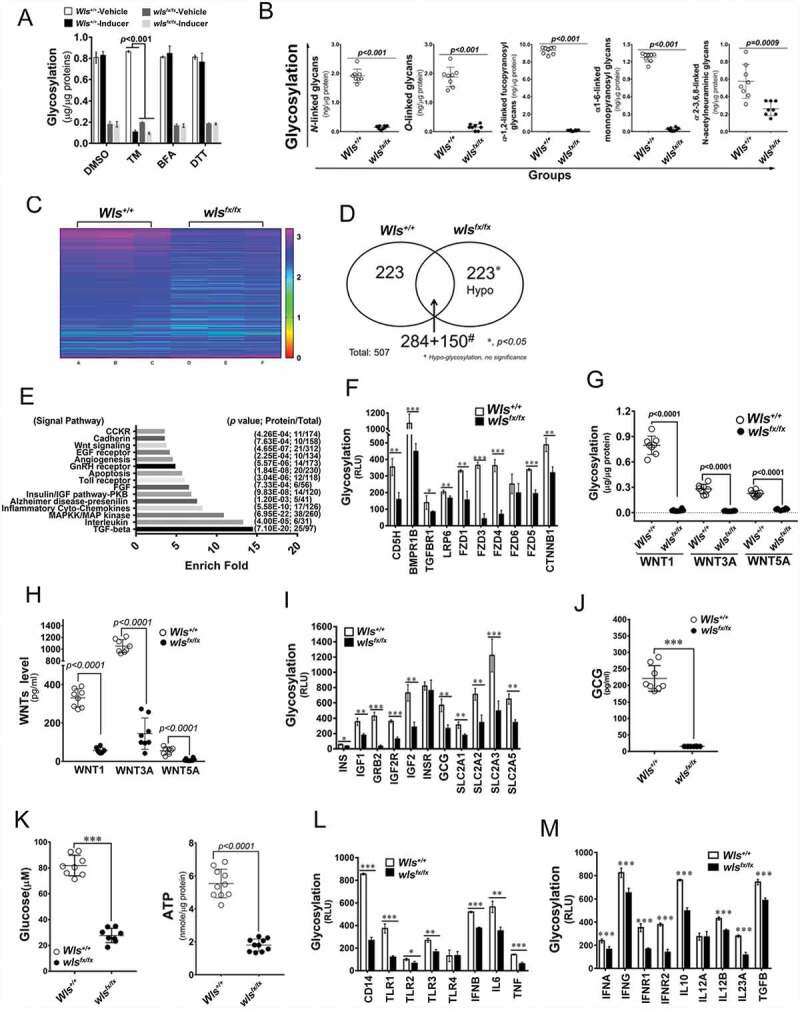
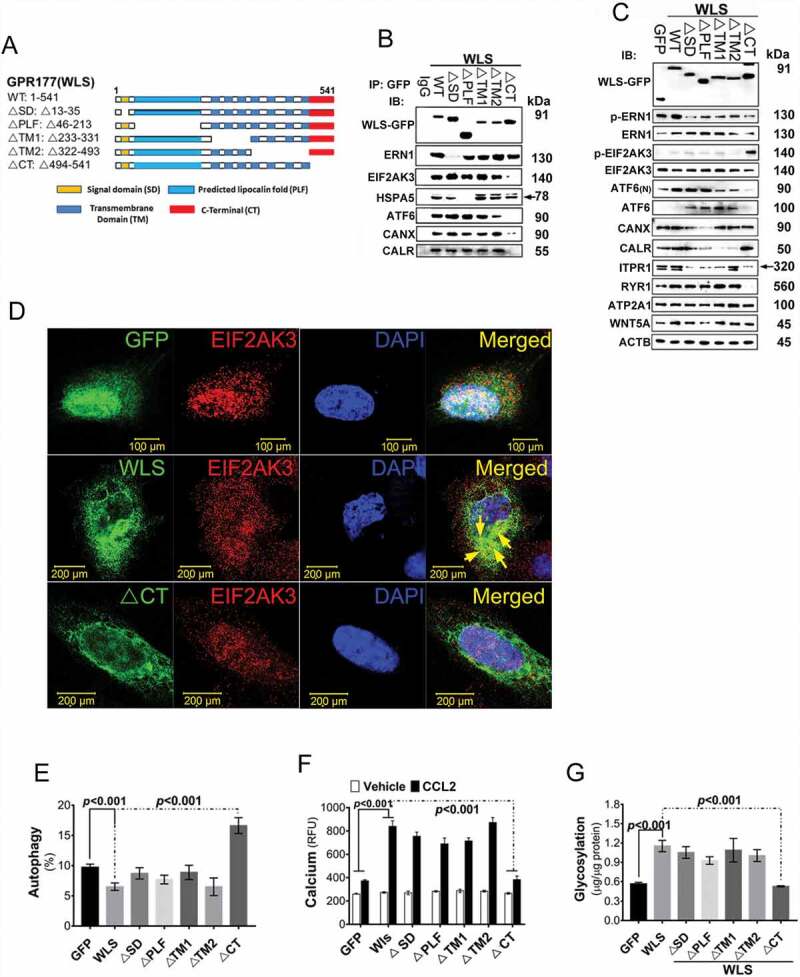
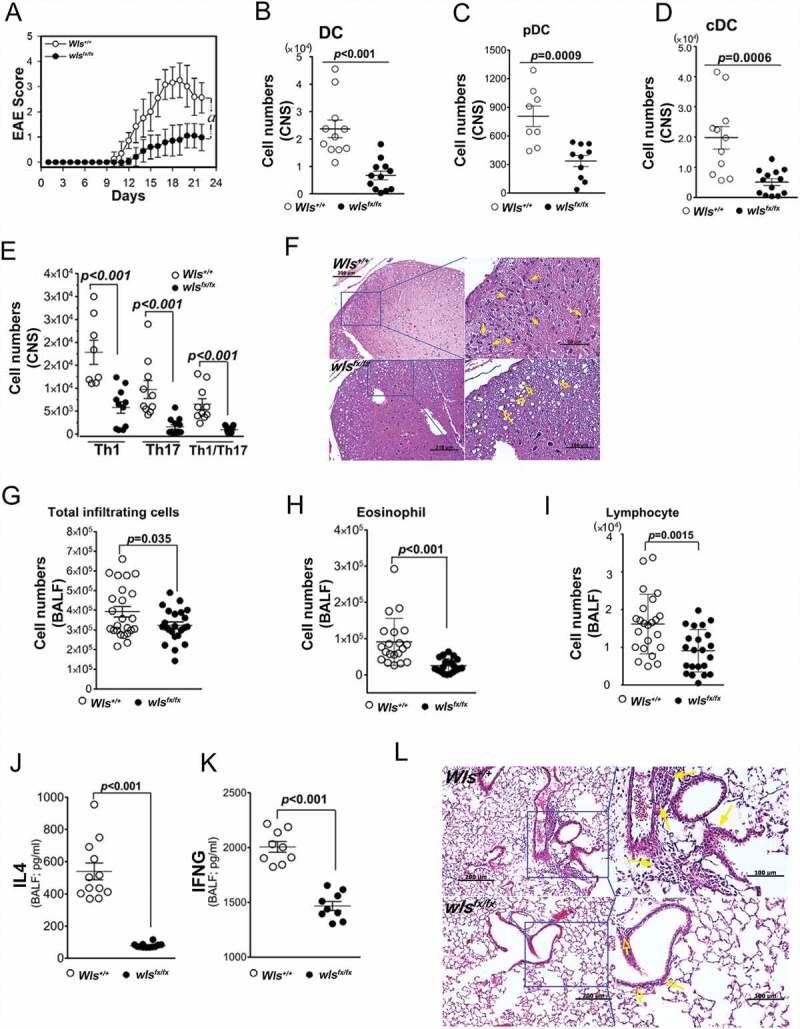
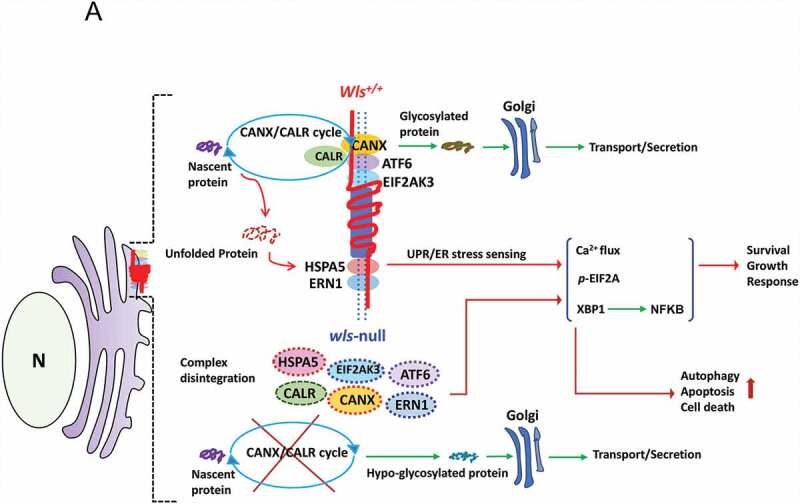
We propose that beyond its role in WNT secretion, WLS/GPR177 (wntless, WNT ligand secretion mediator) acts as an essential regulator controlling protein glycosylation, endoplasmic reticulum (ER) homeostasis, and dendritic cell (DC)-mediated immunity. WLS deficiency in bone marrow-derived DCs (BMDCs) resulted in poor growth and an inability to mount cytokine and T-cell responses in vitro, phenotypes that were irreversible by the addition of exogenous WNTs. In fact, WLS was discovered to integrate a protein complex in N-glycan-dependent and WLS domain-selective manners, comprising ER stress sensors and lectin chaperones. WLS deficiency in BMDCs led to increased ER stress response and macroautophagy/autophagy, decreased calcium efflux from the ER, and the loss of CALR (calreticulin)-CANX (calnexin) cycle, and hence protein hypo-glycosylation. Consequently, DC-specific wls-null mice were unable to develop both Th1-, Th2- and Th17-associated responses in the respective autoimmune and allergic disease models. These results suggest that WLS is a critical chaperone in maintaining ER homeostasis, glycoprotein quality control and calcium dynamics in DCs.Abbreviations: ATF6: activating transcription factor 6; ATG5: autophagy related 5; ATG12: autophagy related 12; ATG16L1: autophagy related 16 like 1; ATP2A1/SERCA1: ATPase sarcoplasmic/endoplasmic reticulum Ca2+ transporting 1; BALF: bronchoalveolar lavage fluid; BFA: brefeldin A; BMDC: bone marrow-derived dendritic cell; CALR: calreticulin; CANX: calnexin; CCL2/MCP-1: C-C motif chemokine ligand 2; CNS: central nervous system; CT: C-terminal domain; DTT: dithiothreitol; DNAJB9/ERDJ4: DnaJ heat shock protein family (Hsp40) member B9; EAE: experimental autoimmune encephalomyelitis; EIF2A/eIF2α: eukaryotic translation initiation factor 2A; EIF2AK3/PERK: eukaryotic translation initiation factor 2 alpha kinase 3; ERN1/IRE1: endoplasmic reticulum (ER) to nucleus signaling 1; GFP: green fluorescent protein; HSPA5/GRP78/BiP: heat shock protein A5; IFNA: interferon alpha; IFNAR1: interferon alpha and beta receptor subunit 1; IFNB: interferon beta; IFNG/INFγ: interferon gamma; IFNGR2: interferon gamma receptor 2; IL6: interleukin 6; IL10: interleukin 10; IL12A: interleukin 12A; IL23A: interleukin 23 subunit alpha; ITGAX/CD11c: integrin subunit alpha X; ITPR1/InsP3R1: inositol 1,4,5-trisphosphate receptor type 1; MAP1LC3B/LC3B: microtubule associated protein 1 light chain 3 beta; OVA: ovalbumin; PIK3C3/VPS34: phosphatidylinositol 3-kinase catalytic subunit type 3; PLF: predicted lipocalin fold; PPP1R15A/GADD34: protein phosphatase 1 regulatory subunit 15A; RYR1/RyanR1: ryanodine receptor 1, skeletal muscle; SD: signal domain; TGFB/TGF-β: transforming growth factor beta family; Th1: T helper cell type 1; Th17: T helper cell type 17; TM: tunicamycin; TNF/TNF-α: tumor necrosis factor; UPR: unfolded protein response; WLS/wntless: WNT ligand secretion mediator.
Keywords: Dendritic cells; er stress; gpr177; protein glycosylation; unfold protein response.
Conflict of interest statement
No potential conflict of interest was reported by the authors.
Figures
References
-
- Banziger C, Soldini D, Schutt C, et al. Wntless, a conserved membrane protein dedicated to the secretion of Wnt proteins from signaling cells. Cell. 2006;125(3):509–522. . - PubMed
-
- Wu BT, Wen SH, Hwang SP, et al. Control of Wnt5b secretion by Wntless modulates chondrogenic cell proliferation through fine-tuning fgf3 expression. J Cell Sci. 2015;128(12):2328–2339. - PubMed
-
- Staal FJ, Luis TC, Tiemessen MM.. WNT signalling in the immune system: WNT is spreading its wings. Nat Rev Immunol. 2008;8(8):581–593. - PubMed
-
- Steinman RM, Granelli-Piperno A, Pope M, et al. The interaction of immunodeficiency viruses with dendritic cells. Curr Top Microbiol Immunol. 2003;276:1–30. - PubMed
-
- Banchereau J, Steinman RM.. Dendritic cells and the control of immunity. Nature. 1998;392(6673):245–252. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous