

MSC-like cells increase ability of monocyte-derived dendritic cells to polarize IL-17-/IL-10-producing T cells via CTLA-4
- PMID: 33855282
- PMCID: PMC8027231
- DOI: 10.1016/j.isci.2021.102312
MSC-like cells increase ability of monocyte-derived dendritic cells to polarize IL-17-/IL-10-producing T cells via CTLA-4
Abstract
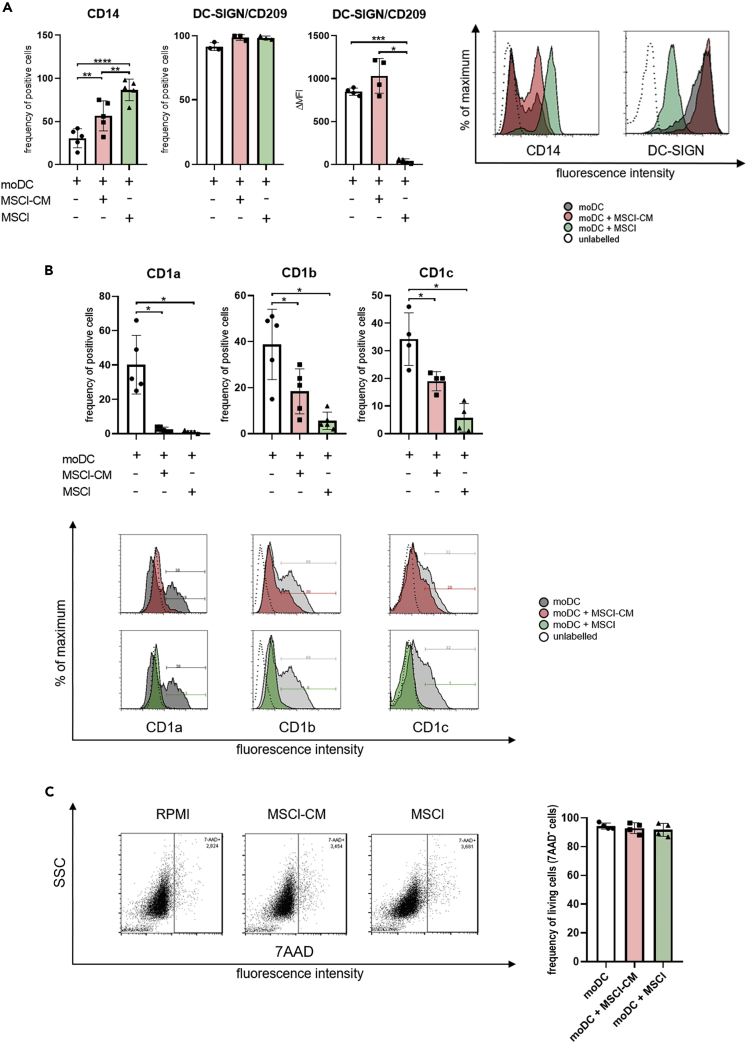
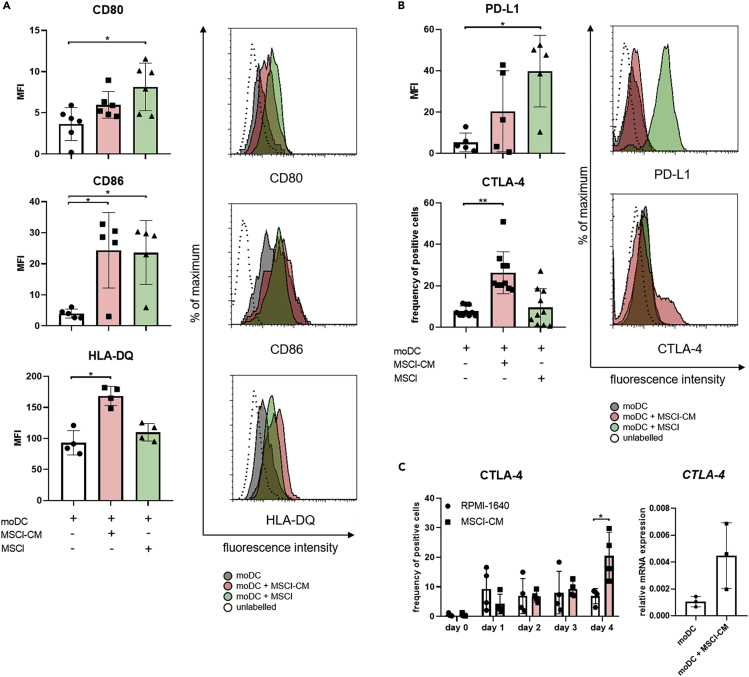
Mesenchymal stromal cell-like (MSCl) cells generated from human embryonic stem cells are considered to be an eligible cell line to model the immunomodulatory behavior of mesenchymal stromal cells (MSCs) in vitro. Dendritic cells (DCs) are essential players in the maintenance and restoration of the sensitive balance between tolerance and immunity. Here, the effects of MSCl cells on the in vitro differentiation of human monocytes into DCs were investigated. MSCl cells promote the differentiation of CTLA-4 expressing DCs via the production of all-trans retinoic acid (ATRA) functioning as a ligand of RARα, a key nuclear receptor in DC development. These semi-matured DCs exhibit an ability to activate allogeneic, naive T cells and polarize them into IL-10 + IL-17 + double-positive T helper cells in a CTLA-4-dependent manner. Mapping the molecular mechanisms of MSC-mediated indirect modulation of DC differentiation may help to expand MSCs' clinical application in cell-free therapies.
Keywords: Components of the Immune System; Immunology; Molecular Biology; Stem Cells Research.
© 2021 The Author(s).
Conflict of interest statement
The authors declare no conflicts of interest related to this research.
Figures




References
-
- Abumaree M.H., Bahattab E., Alsadoun A., Al Dosaimani A., Abomaray F.M., Khatlani T., Kalionis B., El-Muzaini M.F., Alawad A.O., Alaskar A.S. Characterization of the interaction between human decidua parietalis mesenchymal stem/stromal cells and natural killer cells. Stem Cell Res. Ther. 2018;9:102. - PMC - PubMed
-
- Bacskai I., Mazlo A., Kis-Toth K., Szabo A., Panyi G., Sarkadi B., Apati A., Rajnavolgyi E. Mesenchymal stromal cell-like cells set the balance of stimulatory and inhibitory signals in monocyte-derived dendritic cells. Stem Cells Dev. 2015;24:1805–1816. - PubMed
-
- Braza F., Dirou S., Forest V., Sauzeau V., Hassoun D., Chesne J., Cheminant-Muller M.A., Sagan C., Magnan A., Lemarchand P. Mesenchymal stem cells induce suppressive macrophages through phagocytosis in a mouse model of asthma. Stem Cells. 2016;34:1836–1845. - PubMed
-
- Chen P.M., Liu K.J., Hsu P.J., Wei C.F., Bai C.H., Ho L.J., Sytwu H.K., Yen B.L. Induction of immunomodulatory monocytes by human mesenchymal stem cell-derived hepatocyte growth factor through ERK1/2. J. Leukoc. Biol. 2014;96:295–303. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
