

Optimized serum stability and specificity of an αvβ6 integrin-binding peptide for tumor targeting
- PMID: 33857478
- PMCID: PMC8138772
- DOI: 10.1016/j.jbc.2021.100657
Optimized serum stability and specificity of an αvβ6 integrin-binding peptide for tumor targeting
Abstract
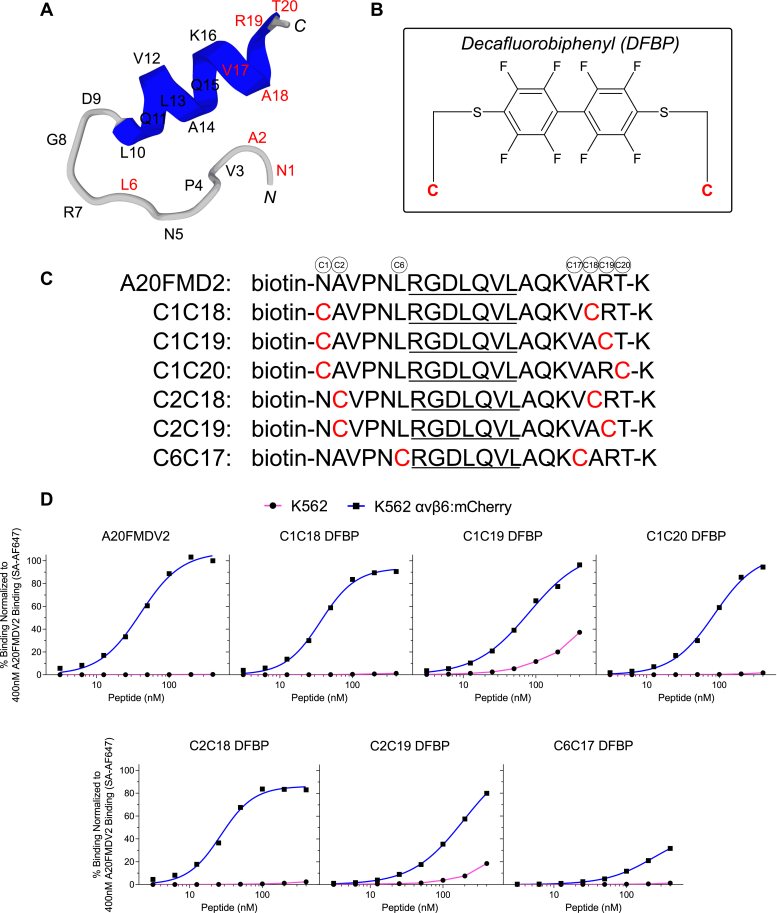
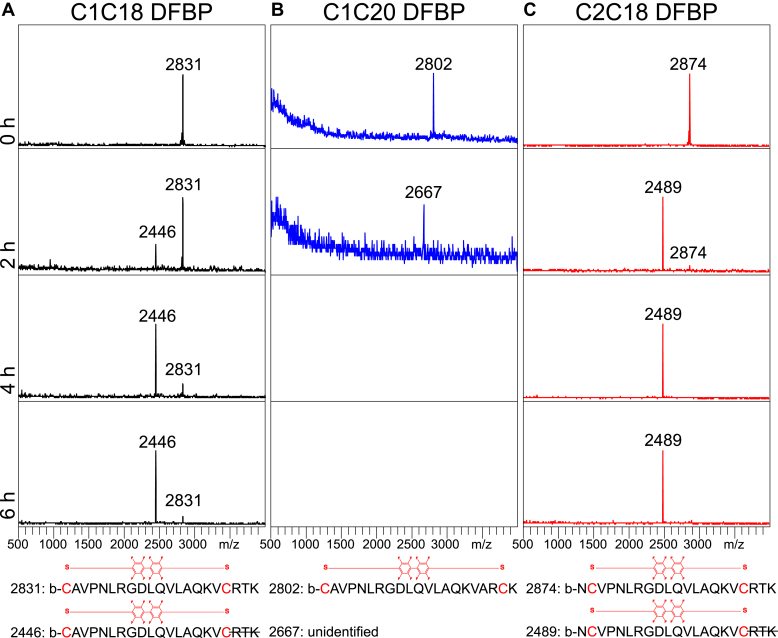
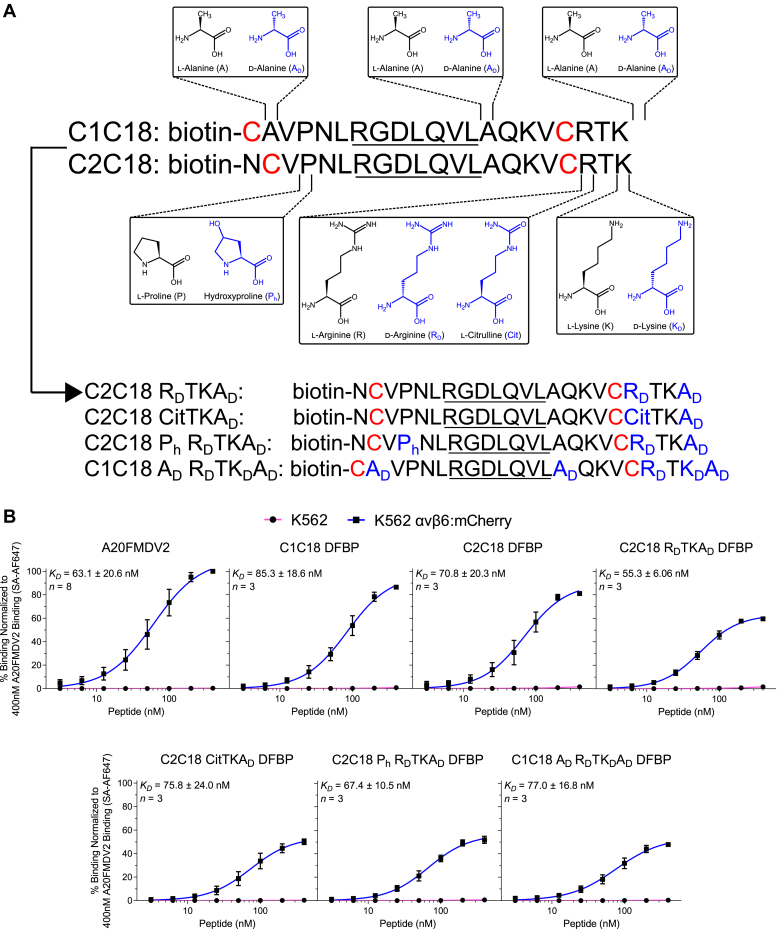
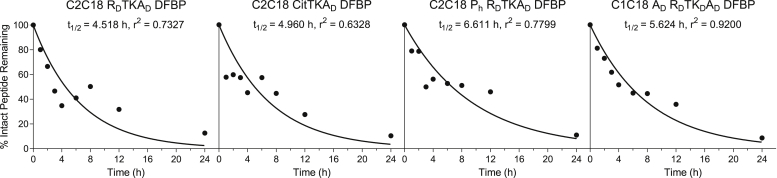
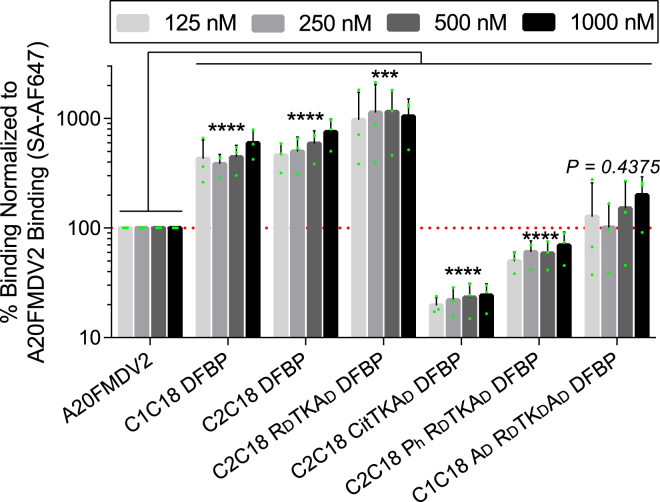
The integrin αvβ6 is an antigen expressed at low levels in healthy tissue but upregulated during tumorigenesis, which makes it a promising target for cancer imaging and therapy. A20FMDV2 is a 20-mer peptide derived from the foot-and-mouth disease virus that exhibits nanomolar and selective affinity for αvβ6 versus other integrins. Despite this selectivity, A20FMDV2 has had limited success in imaging and treating αvβ6+ tumors in vivo because of its poor serum stability. Here, we explore the cyclization and modification of the A20FMDV2 peptide to improve its serum stability without sacrificing its affinity and specificity for αvβ6. Using cysteine amino acid substitutions and cyclization by perfluoroarylation with decafluorobiphenyl, we synthesized six cyclized A20FMDV2 variants and discovered that two retained binding to αvβ6 with modestly improved serum stability. Further d-amino acid substitutions and C-terminal sequence optimization outside the cyclized region greatly prolonged peptide serum stability without reducing binding affinity. While the cyclized A20FMDV2 variants exhibited increased nonspecific integrin binding compared with the original peptide, additional modifications with the non-natural amino acids citrulline, hydroxyproline, and d-alanine were found to restore binding specificity, with some modifications leading to greater αvβ6 integrin selectivity than the original A20FMDV2 peptide. The peptide modifications detailed herein greatly improve the potential of utilizing A20FMDV2 to target αvβ6 in vivo, expanding opportunities for cancer targeting and therapy.
Keywords: A20FMDV2; amino acid substitution; cancer therapy; chemical modification; cyclization; enzymatic stability; integrin; molecular imaging; peptides; αvβ6.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest M. C. J. has interests in Umoja Biopharma and Juno Therapeutics, a Bristol-Myers Squibb company. M. C. J. is a seed investor and holds ownership equity in Umoja, serves as a member of the Umoja Joint Steering Committee, and is a Board Observer of the Umoja Board of Directors. M. C. J. holds patents, some of which are licensed to Umoja Biopharma and Juno Therapeutics. The other authors declare that they have no conflicts of interest with the contents of this article.
Figures

Similar articles
-
Pharmacological Characterization of the αvβ6 Integrin Binding and Internalization Kinetics of the Foot-and-Mouth Disease Virus Derived Peptide A20FMDV2.Pharmacology. 2016;97(3-4):114-25. doi: 10.1159/000443180. Epub 2016 Jan 7. Pharmacology. 2016. PMID: 26734728
-
Use of a peptide derived from foot-and-mouth disease virus for the noninvasive imaging of human cancer: generation and evaluation of 4-[18F]fluorobenzoyl A20FMDV2 for in vivo imaging of integrin alphavbeta6 expression with positron emission tomography.Cancer Res. 2007 Aug 15;67(16):7833-40. doi: 10.1158/0008-5472.CAN-07-1026. Cancer Res. 2007. PMID: 17699789
-
The Effect of Bi-Terminal PEGylation of an Integrin αvβ₆-Targeted ¹⁸F Peptide on Pharmacokinetics and Tumor Uptake.J Nucl Med. 2015 May;56(5):784-90. doi: 10.2967/jnumed.114.150680. Epub 2015 Mar 26. J Nucl Med. 2015. PMID: 25814519 Free PMC article.
-
Harnessing the power of foot-and-mouth-disease virus for targeting integrin alpha-v beta-6 for the therapy of cancer.Expert Opin Drug Discov. 2021 Jul;16(7):737-744. doi: 10.1080/17460441.2021.1878143. Epub 2021 Feb 3. Expert Opin Drug Discov. 2021. PMID: 33533659 Review.
-
111In-Diethylenetriamine pentaacetic acid-NAVPNLRGDLQVLAQKVART.2012 Sep 6 [updated 2012 Nov 29]. In: Molecular Imaging and Contrast Agent Database (MICAD) [Internet]. Bethesda (MD): National Center for Biotechnology Information (US); 2004–2013. 2012 Sep 6 [updated 2012 Nov 29]. In: Molecular Imaging and Contrast Agent Database (MICAD) [Internet]. Bethesda (MD): National Center for Biotechnology Information (US); 2004–2013. PMID: 23193616 Free Books & Documents. Review.
Cited by
-
A comparison of 64Cu-labeled bi-terminally PEGylated A20FMDV2 peptides targeting integrin ανβ6.Oncotarget. 2022 Feb 16;13:360-372. doi: 10.18632/oncotarget.28197. eCollection 2022. Oncotarget. 2022. PMID: 35186193 Free PMC article.
-
Peptide therapeutics in the management of metastatic cancers.RSC Adv. 2022 Aug 2;12(33):21353-21373. doi: 10.1039/d2ra02062a. eCollection 2022 Jul 21. RSC Adv. 2022. PMID: 35975072 Free PMC article. Review.
-
Synthesis and Systematic Study on the Effect of Different PEG Units on Stability of PEGylated, Integrin-αvβ6-Specific A20FMDV2 Analogues in Rat Serum and Human Plasma.Molecules. 2022 Jul 6;27(14):4331. doi: 10.3390/molecules27144331. Molecules. 2022. PMID: 35889207 Free PMC article.
-
Delivery of Theranostic Nanoparticles to Various Cancers by Means of Integrin-Binding Peptides.Int J Mol Sci. 2022 Nov 8;23(22):13735. doi: 10.3390/ijms232213735. Int J Mol Sci. 2022. PMID: 36430214 Free PMC article. Review.
-
Advances in the stability challenges of bioactive peptides and improvement strategies.Curr Res Food Sci. 2022 Nov 5;5:2162-2170. doi: 10.1016/j.crfs.2022.10.031. eCollection 2022. Curr Res Food Sci. 2022. PMID: 36387592 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
