

HLA Ligand Atlas: a benign reference of HLA-presented peptides to improve T-cell-based cancer immunotherapy
- PMID: 33858848
- PMCID: PMC8054196
- DOI: 10.1136/jitc-2020-002071
HLA Ligand Atlas: a benign reference of HLA-presented peptides to improve T-cell-based cancer immunotherapy
Abstract
Background: The human leucocyte antigen (HLA) complex controls adaptive immunity by presenting defined fractions of the intracellular and extracellular protein content to immune cells. Understanding the benign HLA ligand repertoire is a prerequisite to define safe T-cell-based immunotherapies against cancer. Due to the poor availability of benign tissues, if available, normal tissue adjacent to the tumor has been used as a benign surrogate when defining tumor-associated antigens. However, this comparison has proven to be insufficient and even resulted in lethal outcomes. In order to match the tumor immunopeptidome with an equivalent counterpart, we created the HLA Ligand Atlas, the first extensive collection of paired HLA-I and HLA-II immunopeptidomes from 227 benign human tissue samples. This dataset facilitates a balanced comparison between tumor and benign tissues on HLA ligand level.
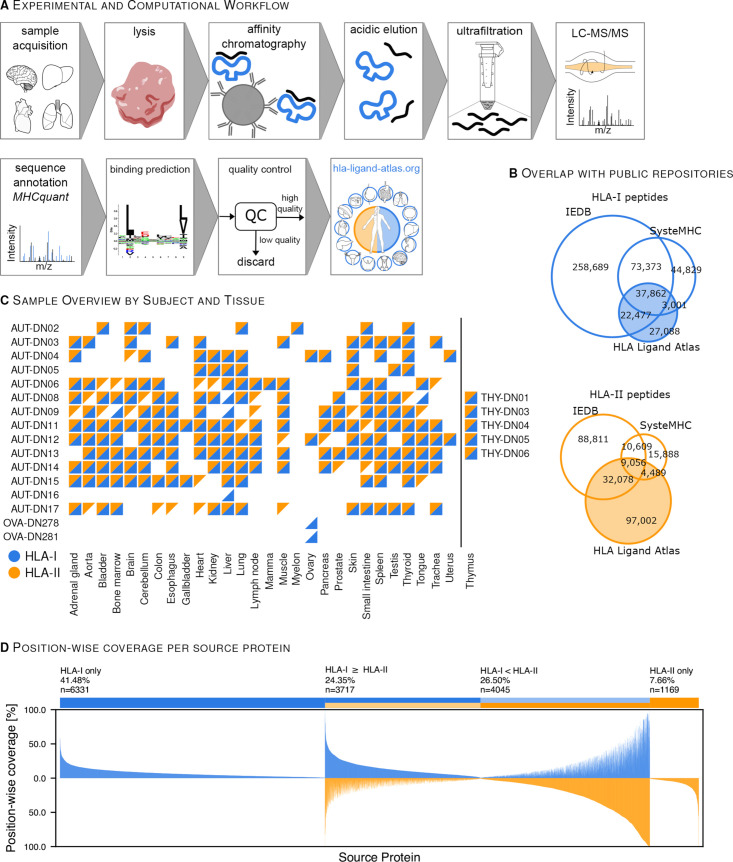
Methods: Human tissue samples were obtained from 16 subjects at autopsy, five thymus samples and two ovary samples originating from living donors. HLA ligands were isolated via immunoaffinity purification and analyzed in over 1200 liquid chromatography mass spectrometry runs. Experimentally and computationally reproducible protocols were employed for data acquisition and processing.
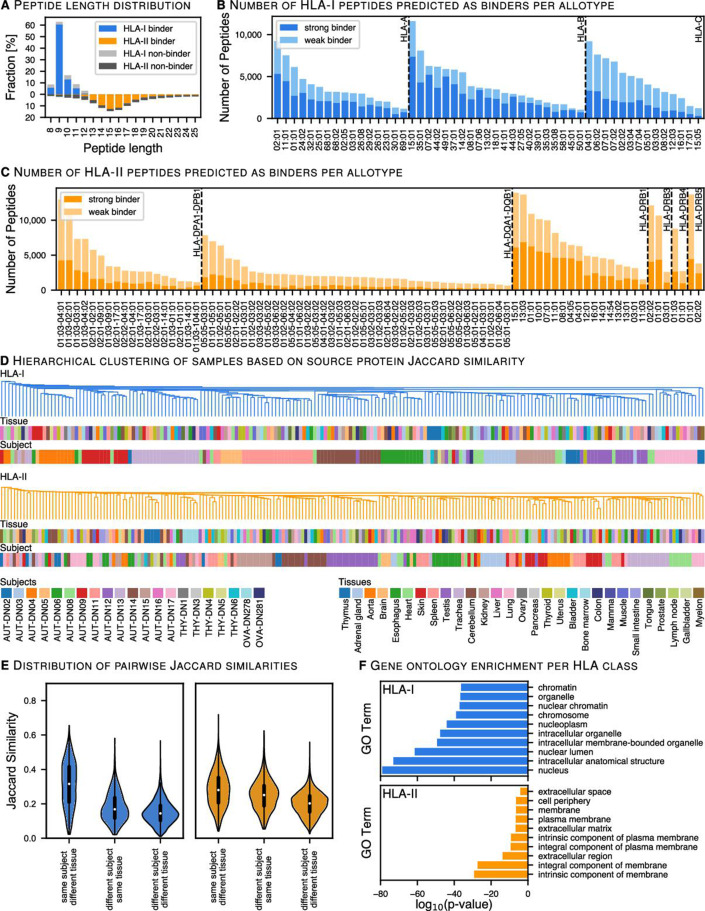
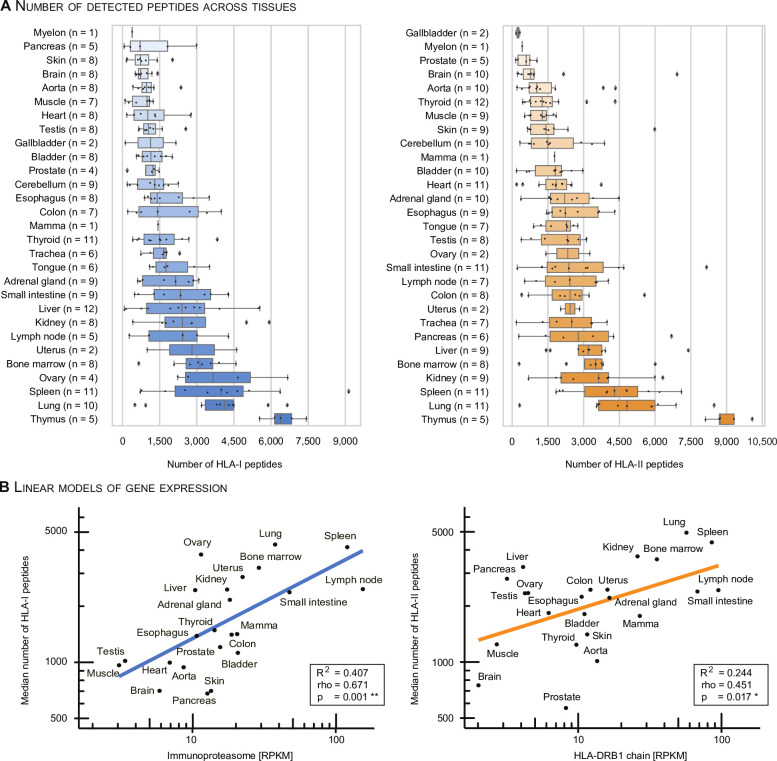
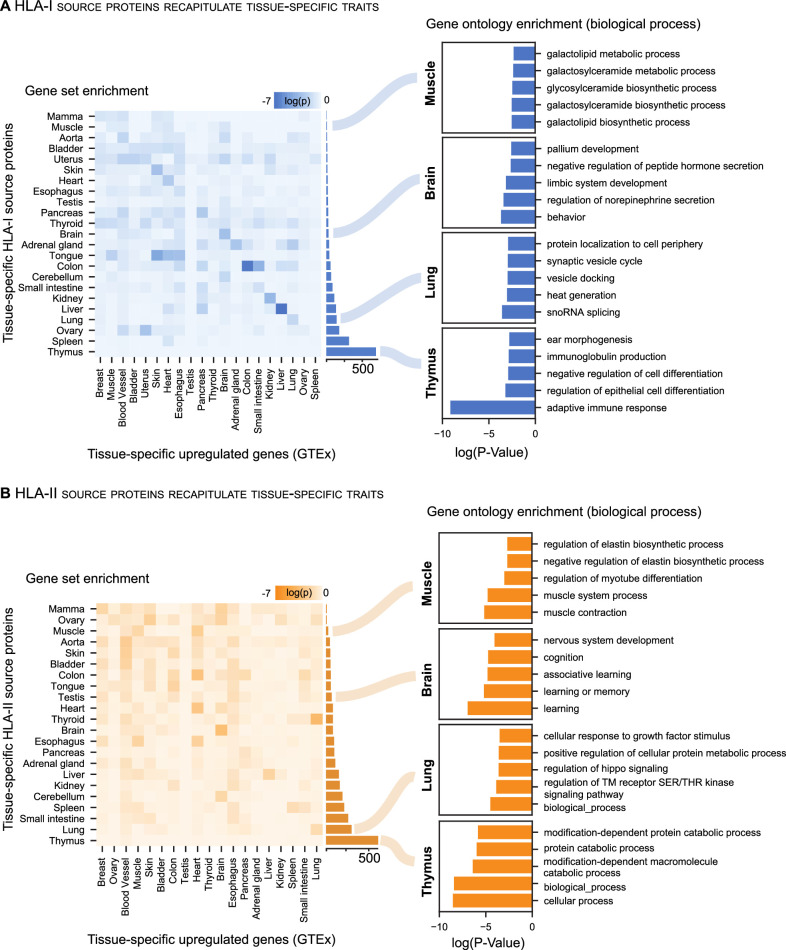
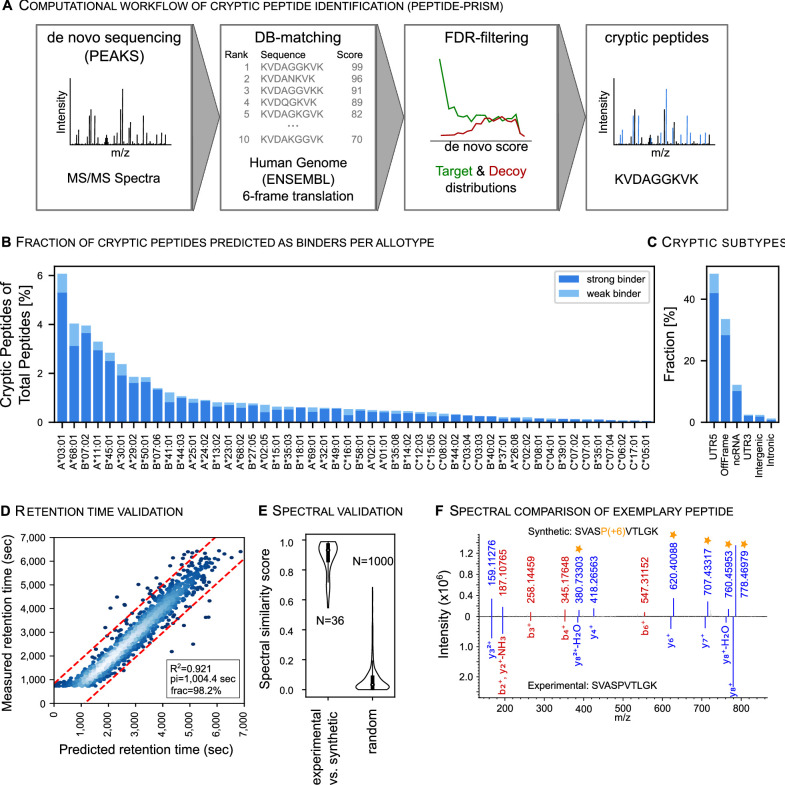
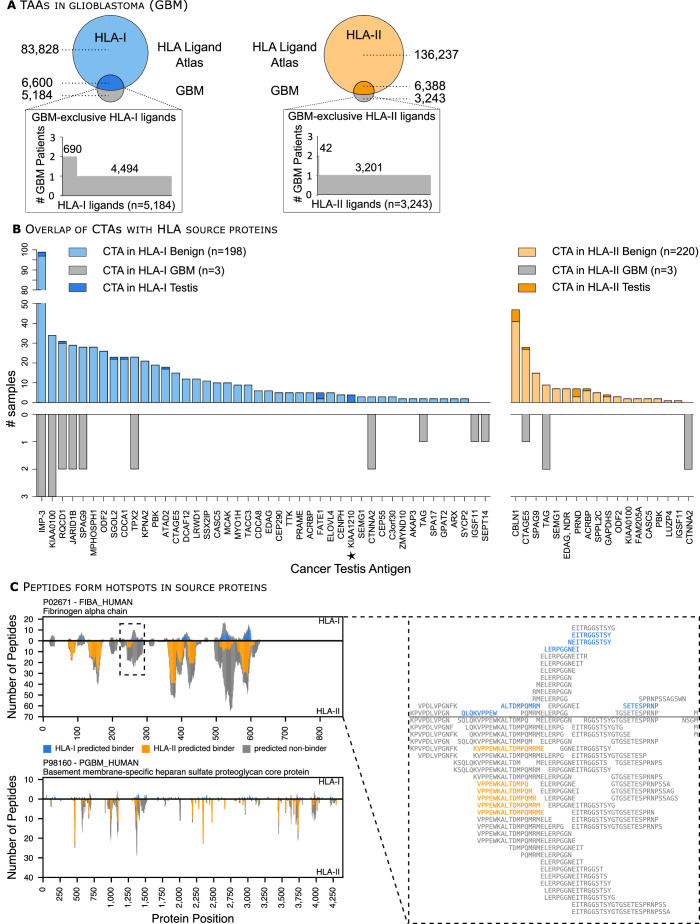
Results: The initial release covers 51 HLA-I and 86 HLA-II allotypes presenting 90,428 HLA-I- and 142,625 HLA-II ligands. The HLA allotypes are representative for the world population. We observe that immunopeptidomes differ considerably between tissues and individuals on source protein and HLA-ligand level. Moreover, we discover 1407 HLA-I ligands from non-canonical genomic regions. Such peptides were previously described in tumors, peripheral blood mononuclear cells (PBMCs), healthy lung tissues and cell lines. In a case study in glioblastoma, we show that potential on-target off-tumor adverse events in immunotherapy can be avoided by comparing tumor immunopeptidomes to the provided multi-tissue reference.
Conclusion: Given that T-cell-based immunotherapies, such as CAR-T cells, affinity-enhanced T cell transfer, cancer vaccines and immune checkpoint inhibition, have significant side effects, the HLA Ligand Atlas is the first step toward defining tumor-associated targets with an improved safety profile. The resource provides insights into basic and applied immune-associated questions in the context of cancer immunotherapy, infection, transplantation, allergy and autoimmunity. It is publicly available and can be browsed in an easy-to-use web interface at https://hla-ligand-atlas.org .
Keywords: adaptive immunity; antigen presentation; antigens; carbohydrate; immunotherapy; translational medical research; tumor-associated.
© Author(s) (or their employer(s)) 2021. Re-use permitted under CC BY. Published by BMJ.
Conflict of interest statement
Competing interests: LB, DJK, ML and LKF are employees of Immatics Biotechnologies. LB, DJK, MWL and SS are inventors of patents owned by Immatics Biotechnologies. MWL has acted as a paid consultant in cancer immunology for Boehringer Ingehlheim Pharma & Co. KG. H-GR is shareholder of Immatics Biotechnologies and Curevac AG.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials