

E2A-regulated epigenetic landscape promotes memory CD8 T cell differentiation
- PMID: 33859041
- PMCID: PMC8072256
- DOI: 10.1073/pnas.2013452118
E2A-regulated epigenetic landscape promotes memory CD8 T cell differentiation
Abstract
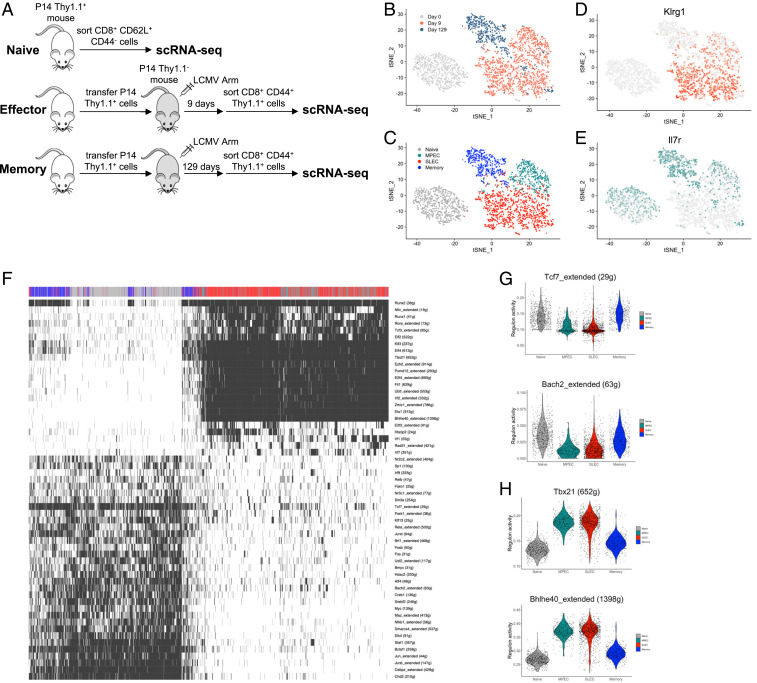
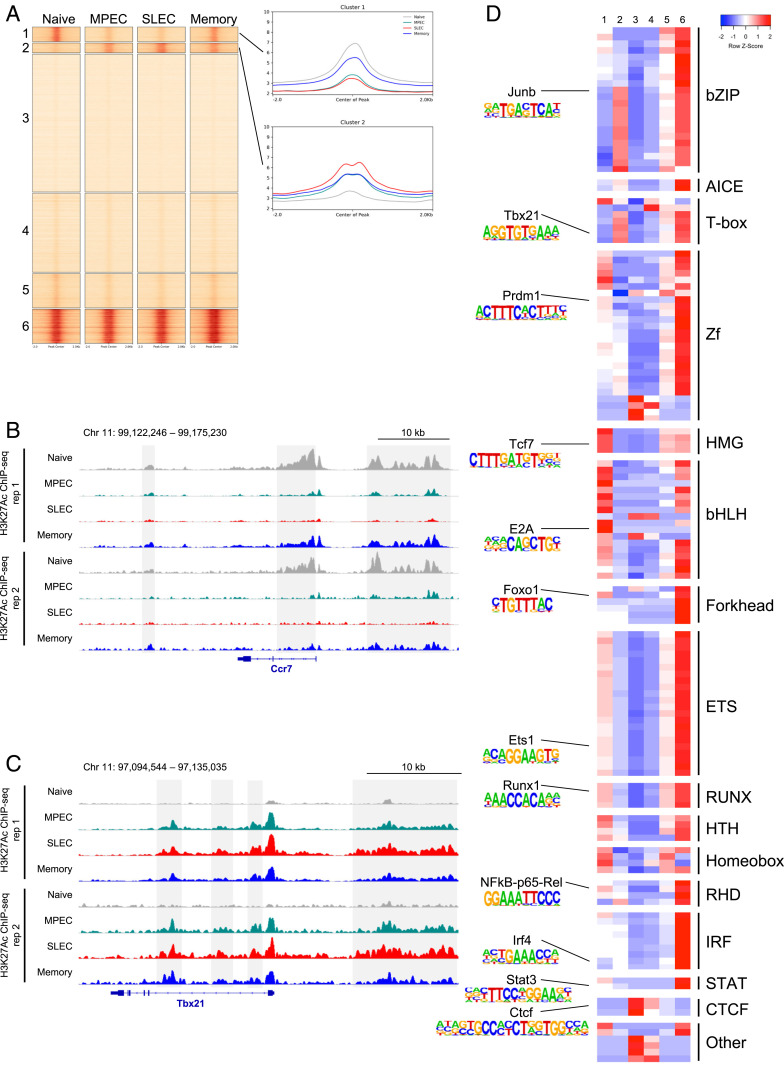
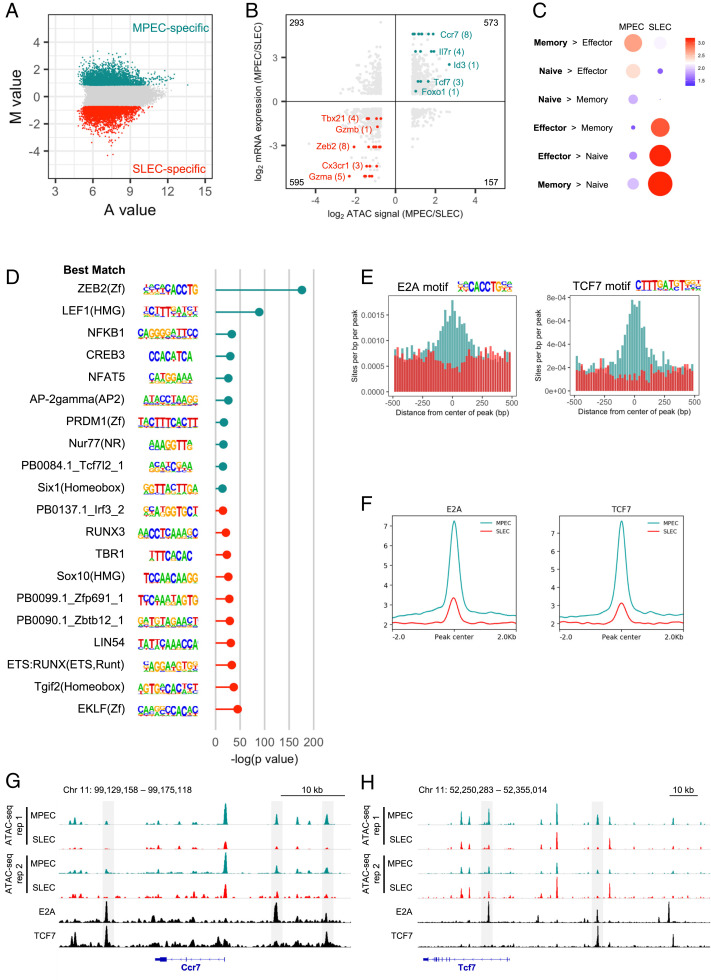
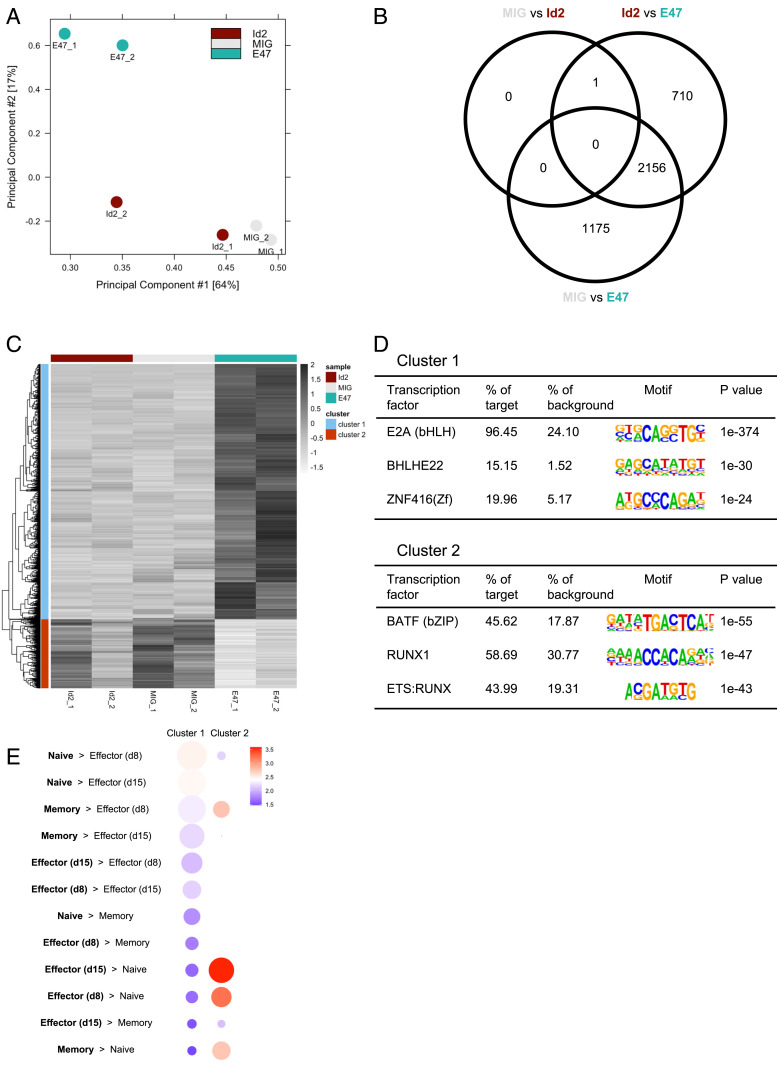
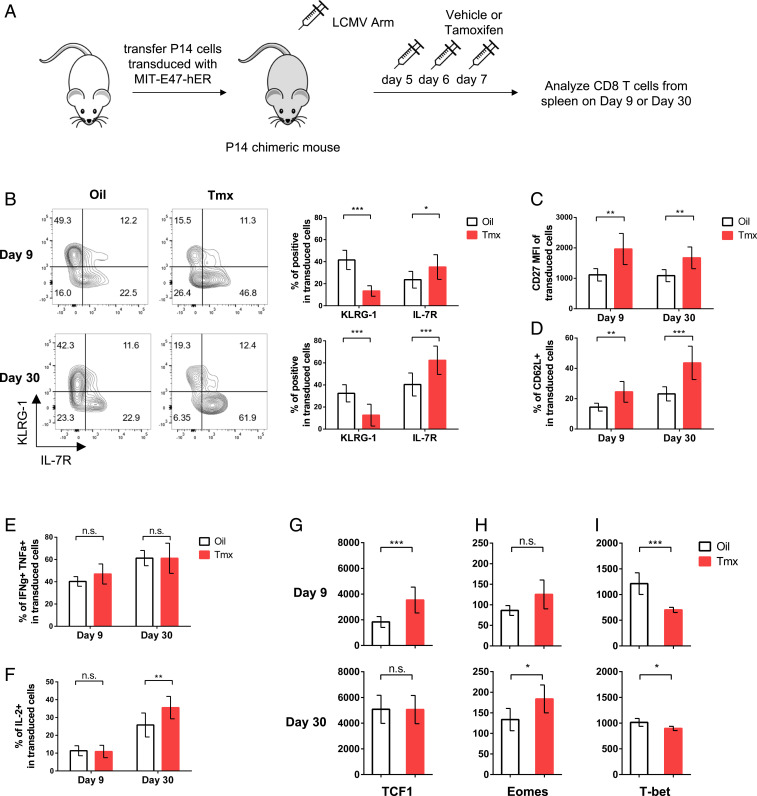
During an acute viral infection, CD8 T cells encounter a myriad of antigenic and inflammatory signals of variable strength, which sets off individual T cells on their own differentiation trajectories. However, the developmental path for each of these cells will ultimately lead to one of only two potential outcomes after clearance of the infection-death or survival and development into memory CD8 T cells. How this cell fate decision is made remains incompletely understood. In this study, we explore the transcriptional changes during effector and memory CD8 T cell differentiation at the single-cell level. Using single-cell, transcriptome-derived gene regulatory network analysis, we identified two main groups of regulons that govern this differentiation process. These regulons function in concert with changes in the enhancer landscape to confer the establishment of the regulatory modules underlying the cell fate decision of CD8 T cells. Furthermore, we found that memory precursor effector cells maintain chromatin accessibility at enhancers for key memory-related genes and that these enhancers are highly enriched for E2A binding sites. Finally, we show that E2A directly regulates accessibility of enhancers of many memory-related genes and that its overexpression increases the frequency of memory precursor effector cells and accelerates memory cell formation while decreasing the frequency of short-lived effector cells. Overall, our results suggest that effector and memory CD8 T cell differentiation is largely regulated by two transcriptional circuits, with E2A serving as an important epigenetic regulator of the memory circuit.
Keywords: CD8 T cell; LCMV; T cell memory; epigenetics; scRNA-seq.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Kaech S. M., et al., Selective expression of the interleukin 7 receptor identifies effector CD8 T cells that give rise to long-lived memory cells. Nat. Immunol. 4, 1191–1198 (2003). - PubMed
-
- Schluns K. S., Kieper W. C., Jameson S. C., Lefrançois L., Interleukin-7 mediates the homeostasis of naïve and memory CD8 T cells in vivo. Nat. Immunol. 1, 426–432 (2000). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
