

Selenoprotein W ensures physiological bone remodeling by preventing hyperactivity of osteoclasts
- PMID: 33859201
- PMCID: PMC8050258
- DOI: 10.1038/s41467-021-22565-7
Selenoprotein W ensures physiological bone remodeling by preventing hyperactivity of osteoclasts
Abstract
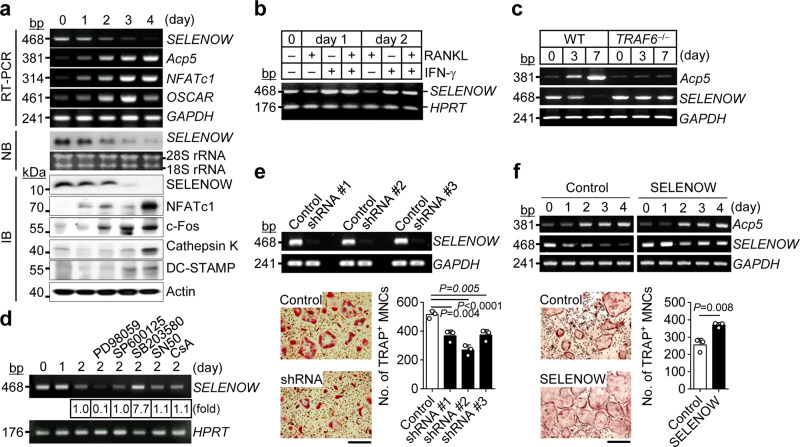
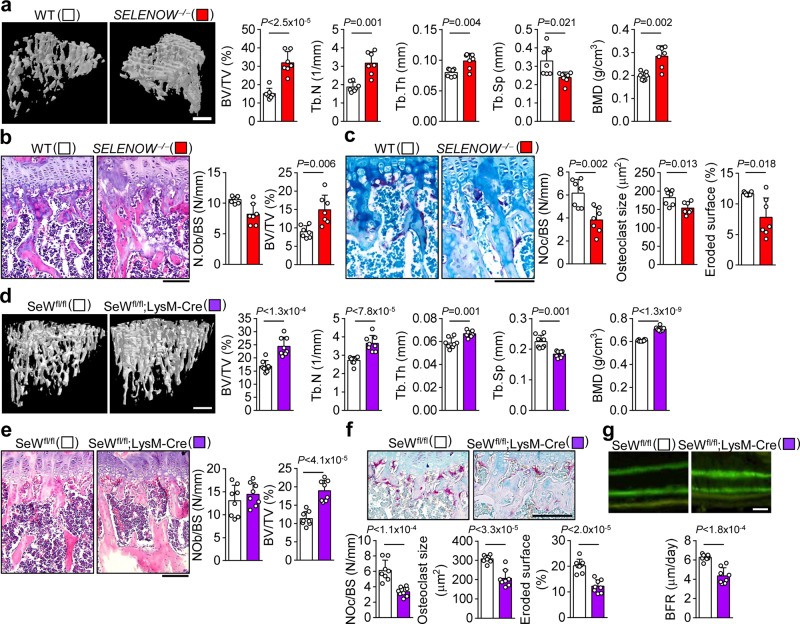
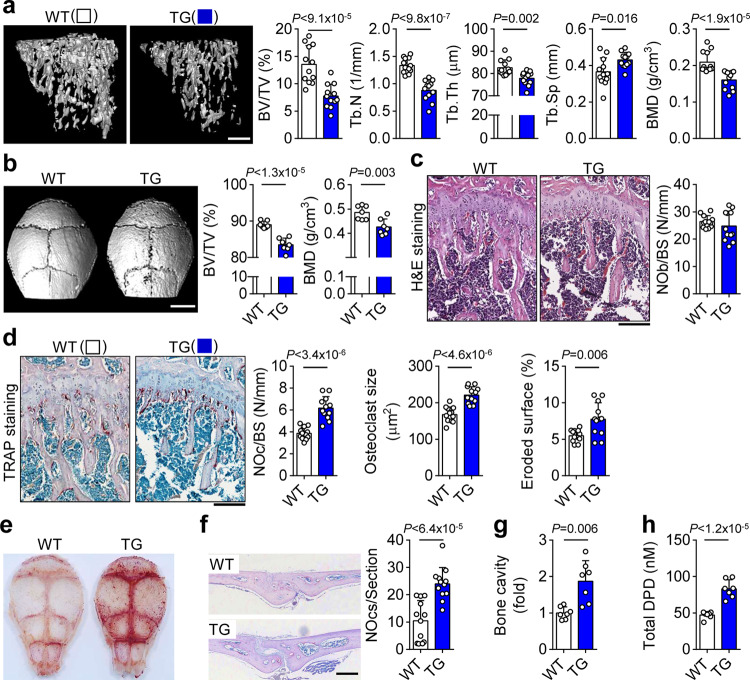
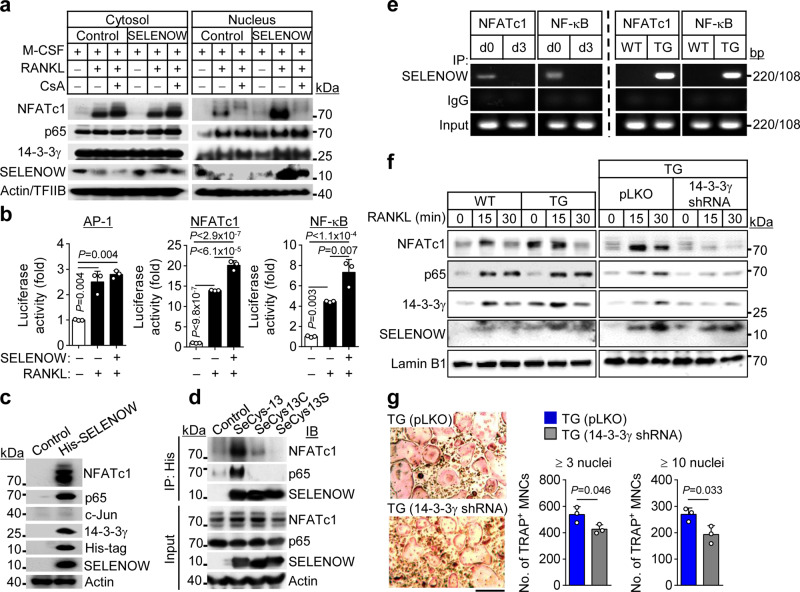
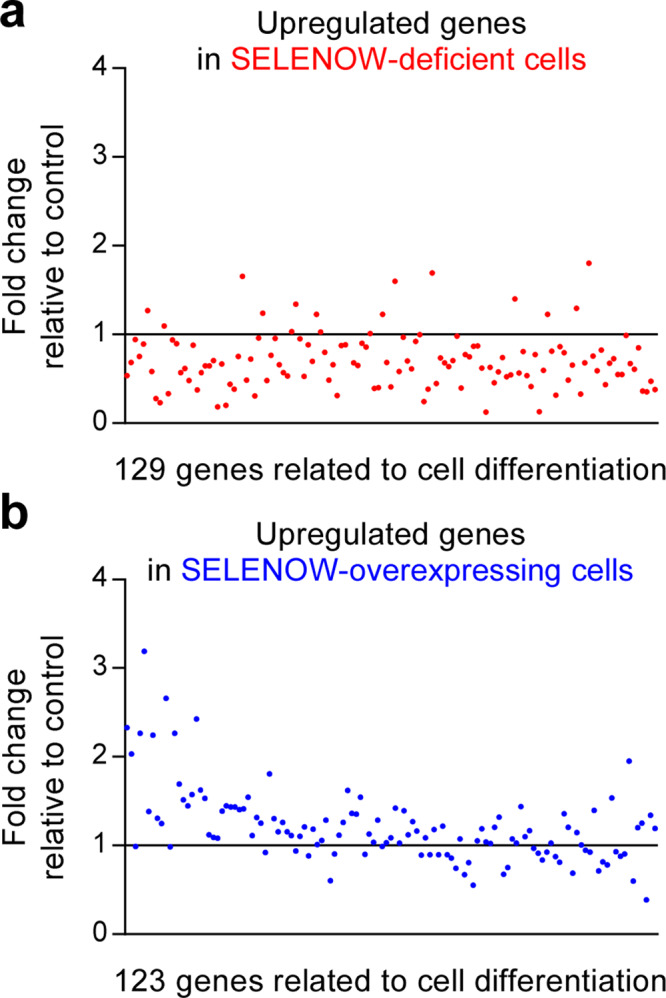
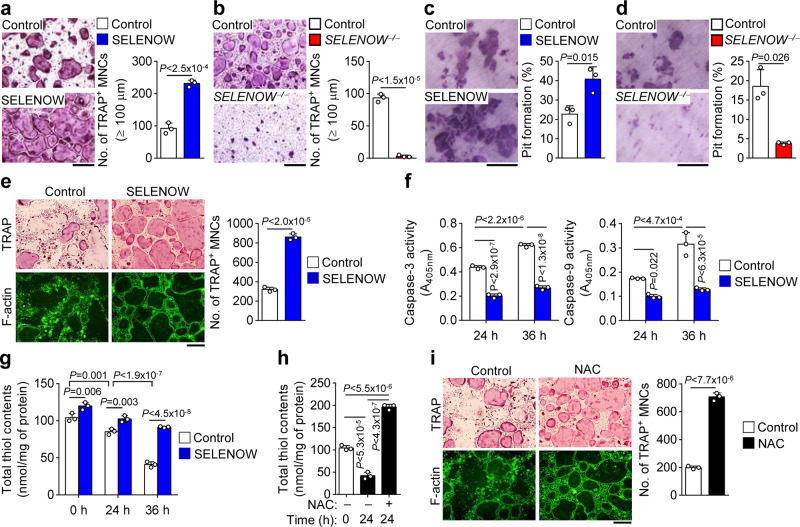
Selenoproteins containing selenium in the form of selenocysteine are critical for bone remodeling. However, their underlying mechanism of action is not fully understood. Herein, we report the identification of selenoprotein W (SELENOW) through large-scale mRNA profiling of receptor activator of nuclear factor (NF)-κΒ ligand (RANKL)-induced osteoclast differentiation, as a protein that is downregulated via RANKL/RANK/tumour necrosis factor receptor-associated factor 6/p38 signaling. RNA-sequencing analysis revealed that SELENOW regulates osteoclastogenic genes. SELENOW overexpression enhances osteoclastogenesis in vitro via nuclear translocation of NF-κB and nuclear factor of activated T-cells cytoplasmic 1 mediated by 14-3-3γ, whereas its deficiency suppresses osteoclast formation. SELENOW-deficient and SELENOW-overexpressing mice exhibit high bone mass phenotype and osteoporosis, respectively. Ectopic SELENOW expression stimulates cell-cell fusion critical for osteoclast maturation as well as bone resorption. Thus, RANKL-dependent repression of SELENOW regulates osteoclast differentiation and blocks osteoporosis caused by overactive osteoclasts. These findings demonstrate a biological link between selenium and bone metabolism.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Hancock, E. J., Ang, J., Papachristodoulou, A. & Stan, G. B. The interplay between feedback and buffering in cellular homeostasis. Cell Syst. 5, 498–508.e23 (2017). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
