

Interleukin-37 regulates innate immune signaling in human and mouse colonic organoids
- PMID: 33859245
- PMCID: PMC8050237
- DOI: 10.1038/s41598-021-87592-2
Interleukin-37 regulates innate immune signaling in human and mouse colonic organoids
Abstract
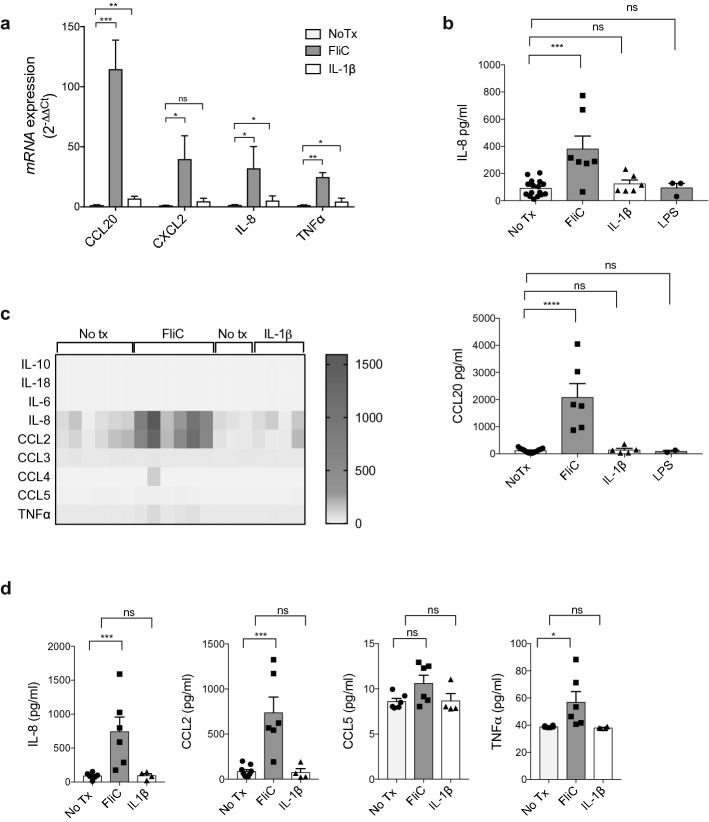
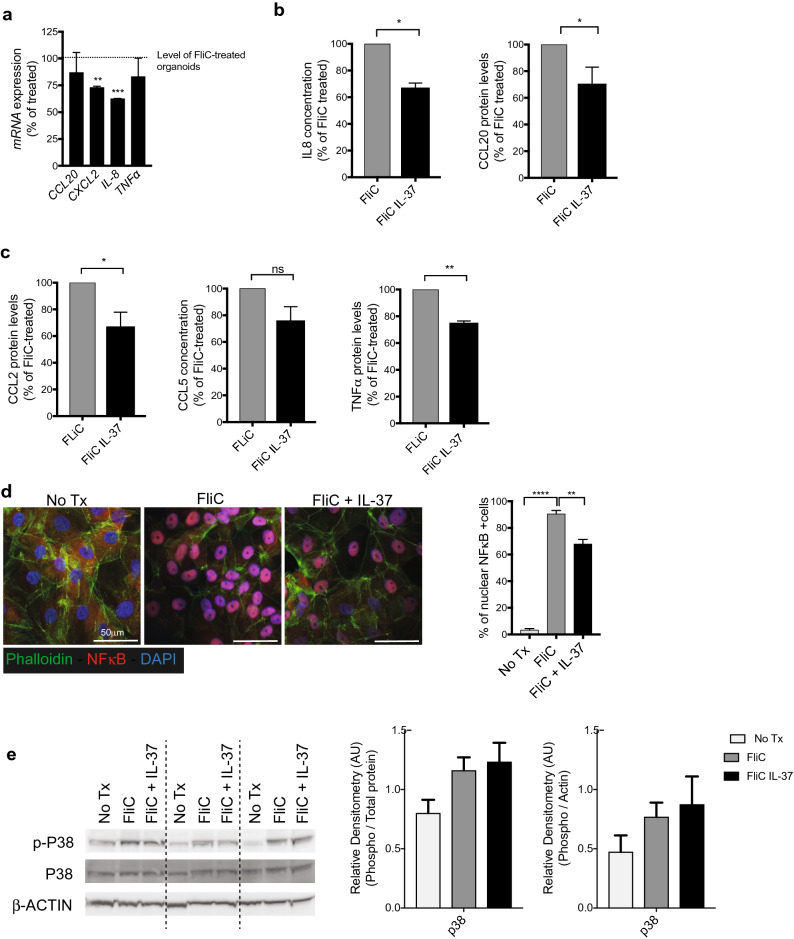
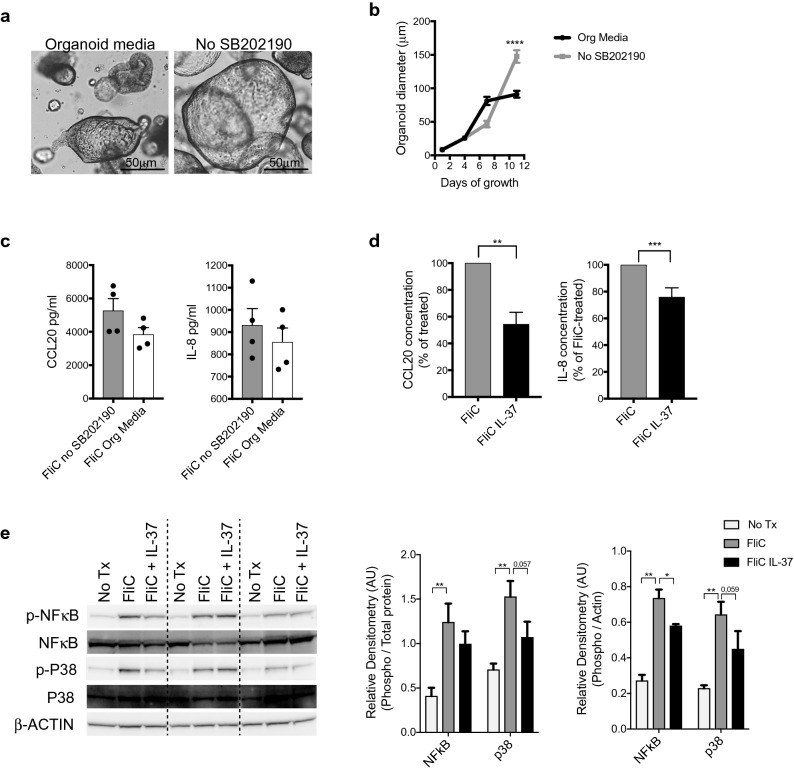
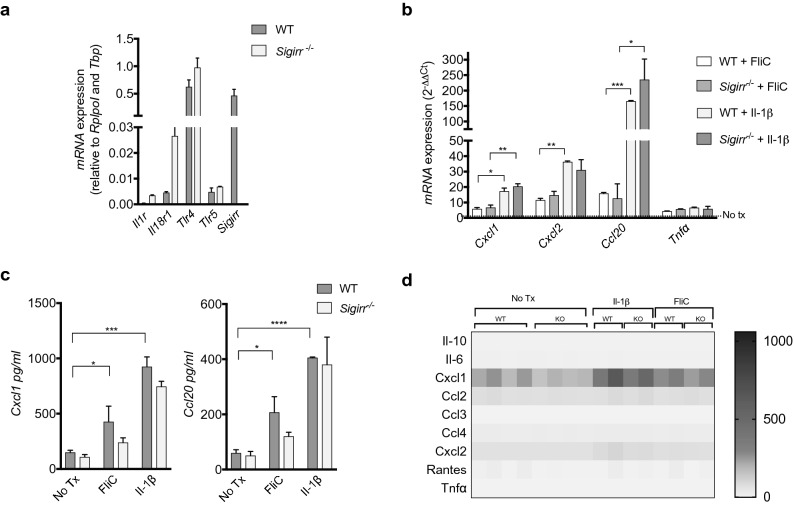
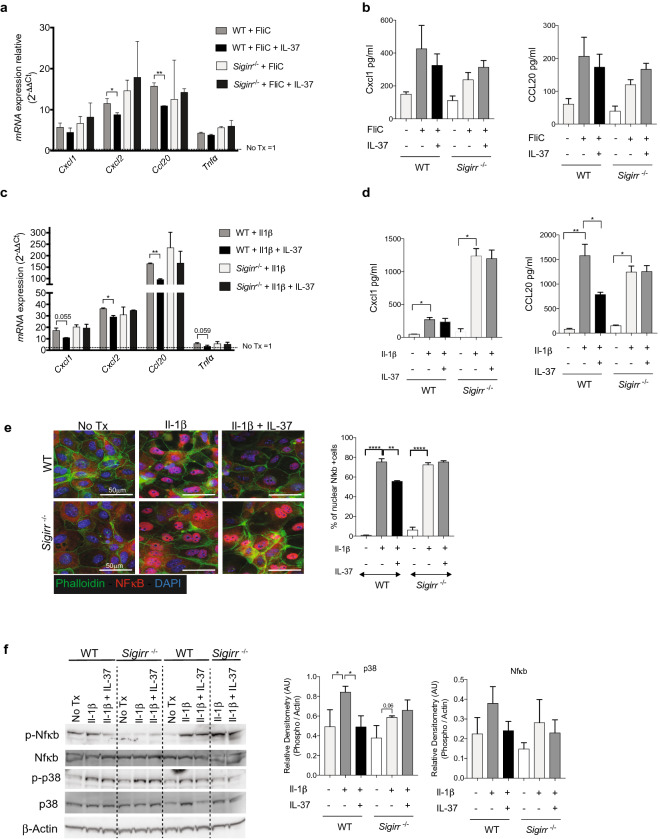
Intestinal epithelial cells (IEC) reside in close proximity to the gut microbiota and are hypo-responsive to bacterial products, likely to prevent maladaptive inflammatory responses. This is in part due to their strong expression of Single Ig IL-1 related receptor (SIGIRR), a negative regulator of interleukin (IL)-1 and toll-like receptor signaling. IL-37 is an anti-inflammatory cytokine that inhibits innate signaling in diverse cells by signaling through SIGIRR. Despite the strong expression of SIGIRR by IEC, few studies have examined whether IL-37 can suppress their innate immune signaling. We characterized innate immune responses of human and murine colonoids to bacteria (FliC, LPS) and host (IL-1β) products and the role of IL-37/SIGIRR in regulating these responses. We demonstrated that human colonoids responded only to FliC, but not to LPS or IL-1β. While colonoids derived from different donors displayed significant inter-individual variability in the magnitude of their innate responses to FliC stimulation, all colonoids released a variety of chemokines. Interestingly, IL-37 attenuated these responses through inhibition of p38 and NFκB signaling pathways. We determined that this suppression by IL-37 was SIGIRR dependent, in murine organoids. Along with species-specific differences in IEC innate responses, we show that IL-37 can promote IEC hypo-responsiveness by suppressing inflammatory signaling.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Frantz AL, et al. Targeted deletion of MyD88 in intestinal epithelial cells results in compromised antibacterial immunity associated with downregulation of polymeric immunoglobulin receptor, mucin-2, and antibacterial peptides. Mucosal Immunol. 2012;5:501–512. doi: 10.1038/mi.2012.23. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
