

Generation of recombinant hyperimmune globulins from diverse B-cell repertoires
- PMID: 33859400
- PMCID: PMC8355030
- DOI: 10.1038/s41587-021-00894-8
Generation of recombinant hyperimmune globulins from diverse B-cell repertoires
Abstract
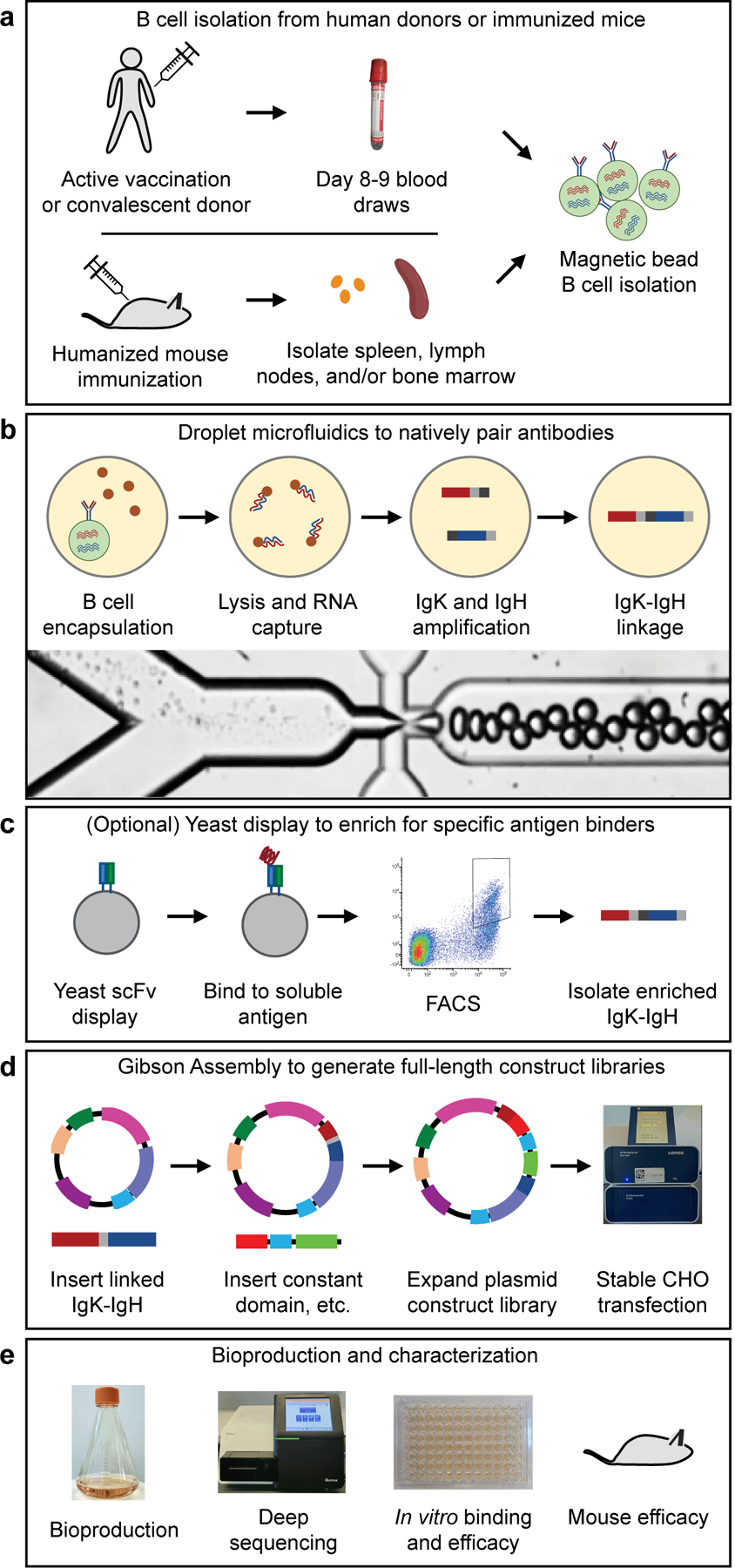
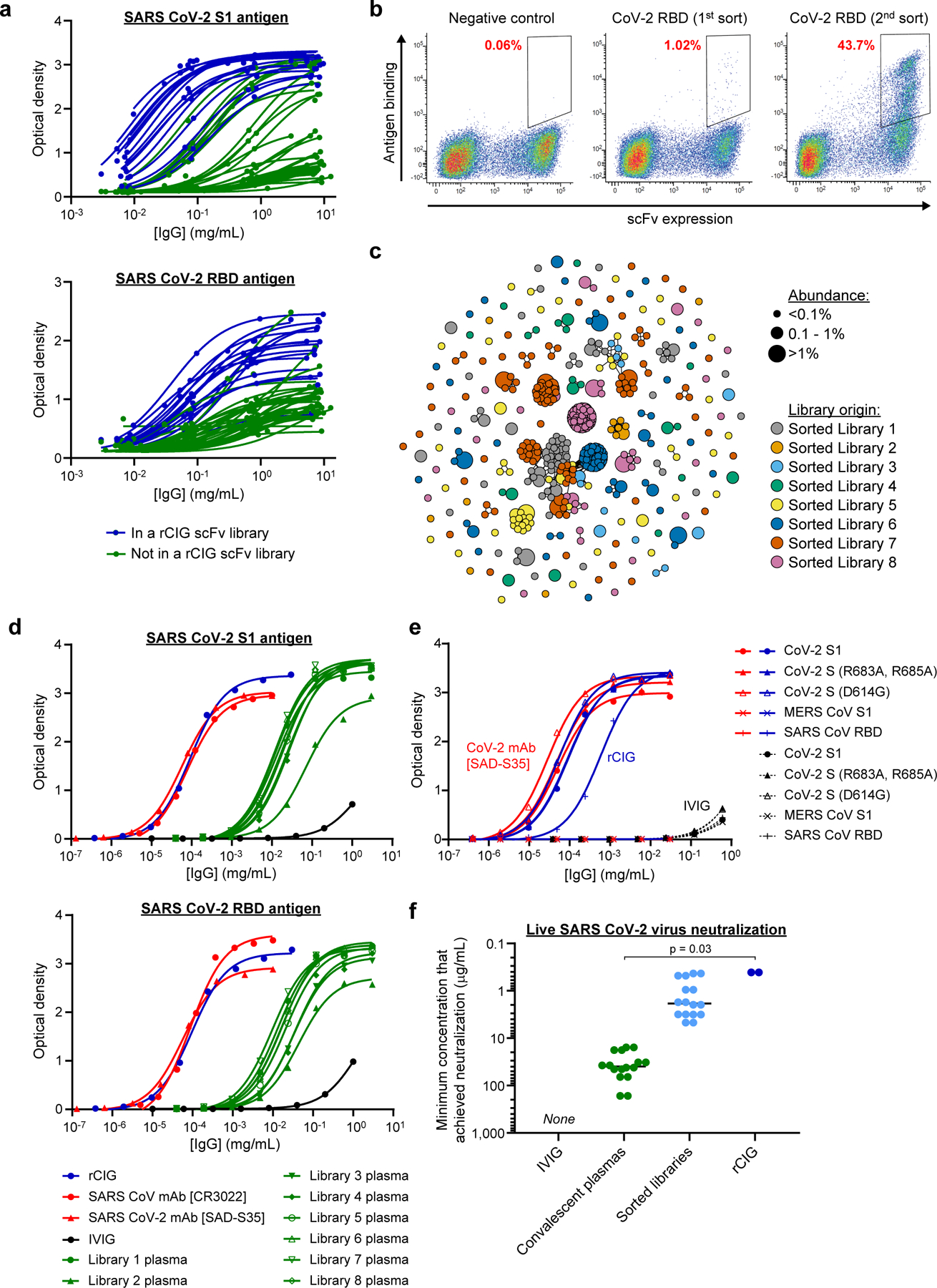
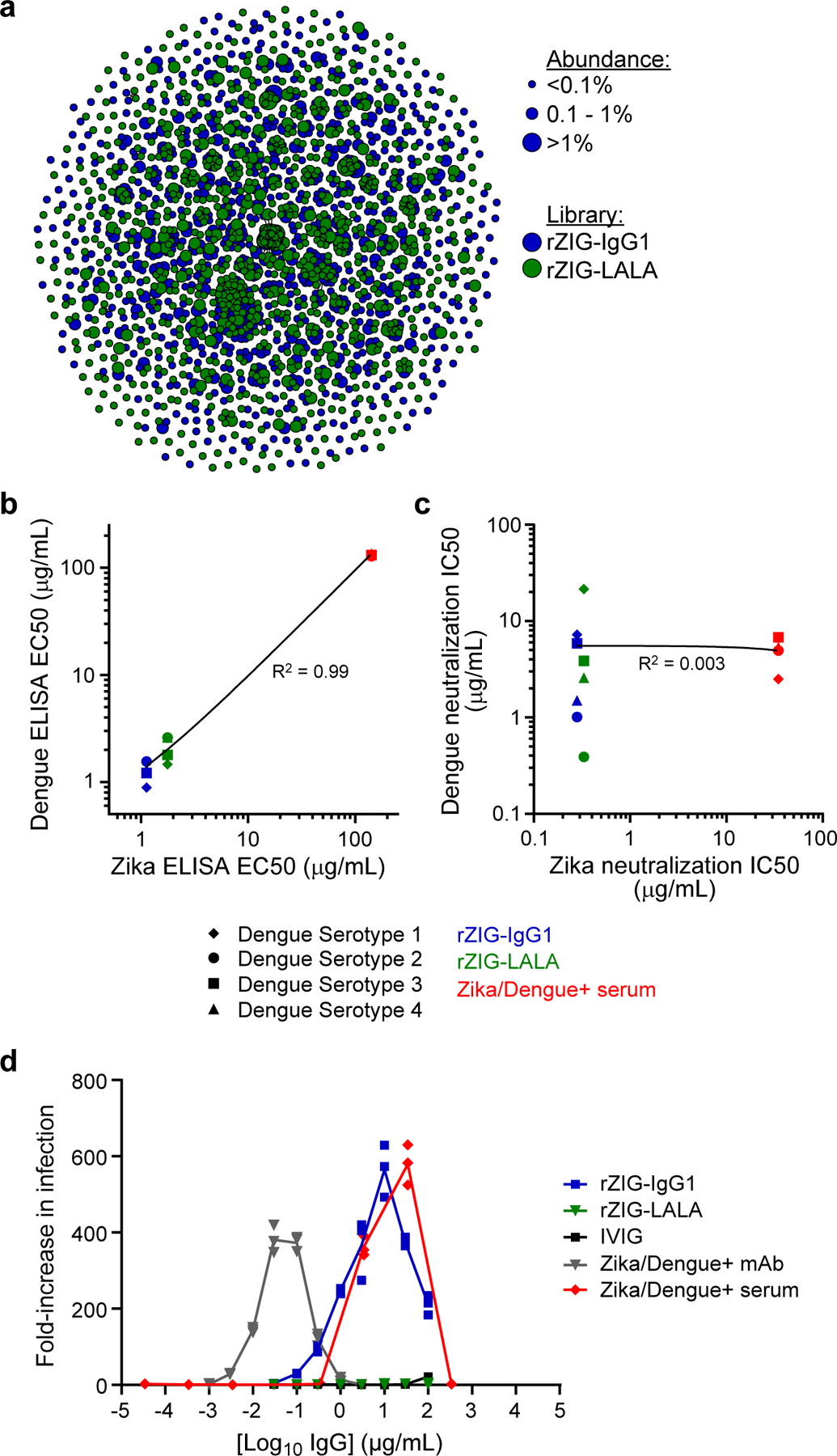
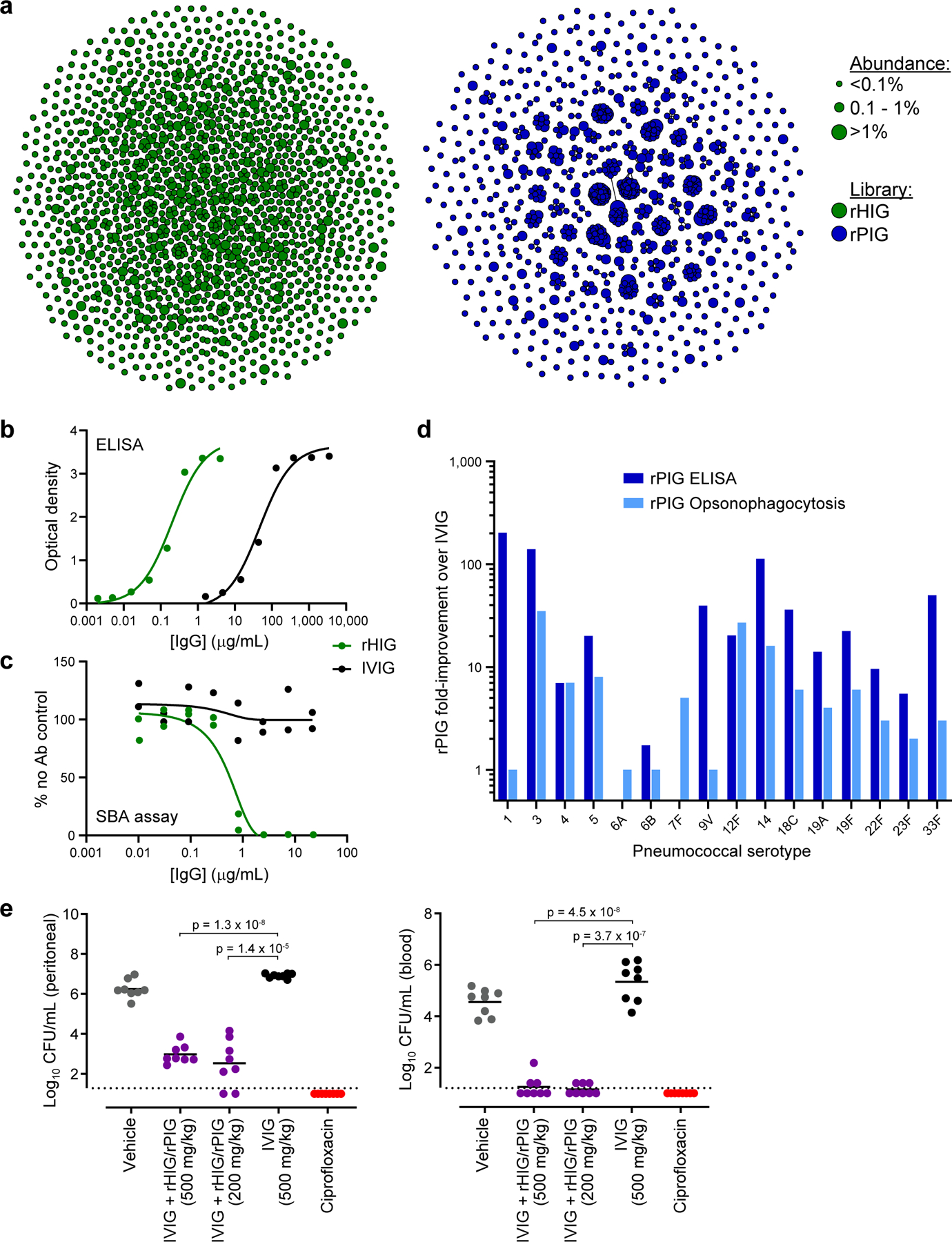
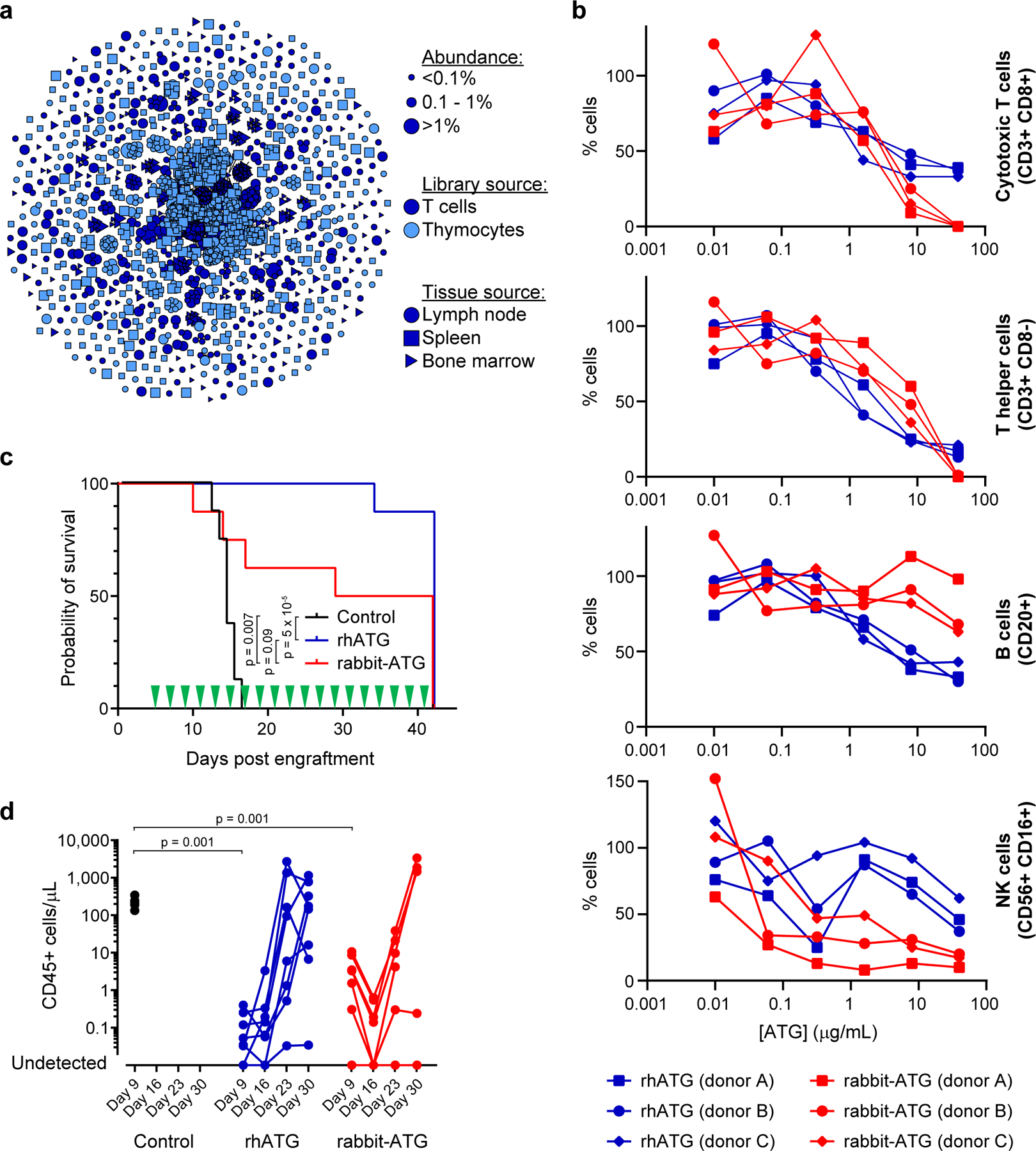
Plasma-derived polyclonal antibody therapeutics, such as intravenous immunoglobulin, have multiple drawbacks, including low potency, impurities, insufficient supply and batch-to-batch variation. Here we describe a microfluidics and molecular genomics strategy for capturing diverse mammalian antibody repertoires to create recombinant multivalent hyperimmune globulins. Our method generates of diverse mixtures of thousands of recombinant antibodies, enriched for specificity and activity against therapeutic targets. Each hyperimmune globulin product comprised thousands to tens of thousands of antibodies derived from convalescent or vaccinated human donors or from immunized mice. Using this approach, we generated hyperimmune globulins with potent neutralizing activity against severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) in under 3 months, Fc-engineered hyperimmune globulins specific for Zika virus that lacked antibody-dependent enhancement of disease, and hyperimmune globulins specific for lung pathogens present in patients with primary immune deficiency. To address the limitations of rabbit-derived anti-thymocyte globulin, we generated a recombinant human version and demonstrated its efficacy in mice against graft-versus-host disease.
© 2021. The Author(s), under exclusive licence to Springer Nature America, Inc.
Figures
References
-
- Beasley RP, Hwang LY, Stevens CE, et al. Efficacy of hepatitis B immune globulin for prevention of perinatal transmission of the hepatitis B virus carrier state: final report of a randomized double-blind, placebo-controlled trial. Hepatology 1983;3(2):135–141. doi: 10.1002/hep.1840030201 - DOI - PubMed
-
- Gaber AO, First MR, Tesi RJ, et al. Results of the double-blind, randomized, multicenter, phase III clinical trial of Thymoglobulin versus Atgam in the treatment of acute graft rejection episodes after renal transplantation. Transplantation 1998;66(1):29–37. doi: 10.1097/00007890-199807150-00005 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
