

The nucleolus as a polarized coaxial cable in which the rDNA axis is surrounded by dynamic subunit-specific phases
- PMID: 33862007
- PMCID: PMC8222187
- DOI: 10.1016/j.cub.2021.03.041
The nucleolus as a polarized coaxial cable in which the rDNA axis is surrounded by dynamic subunit-specific phases
Abstract
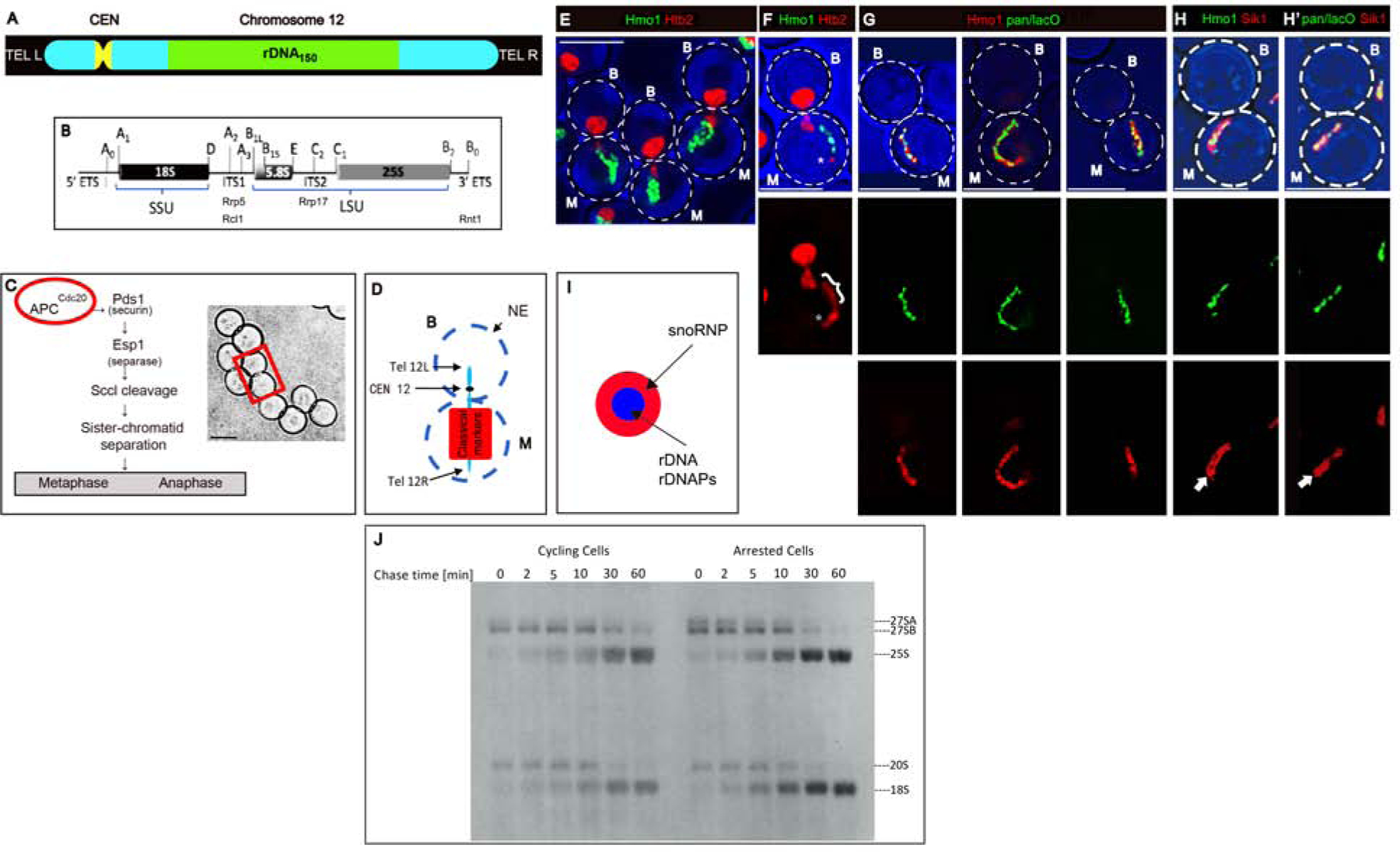
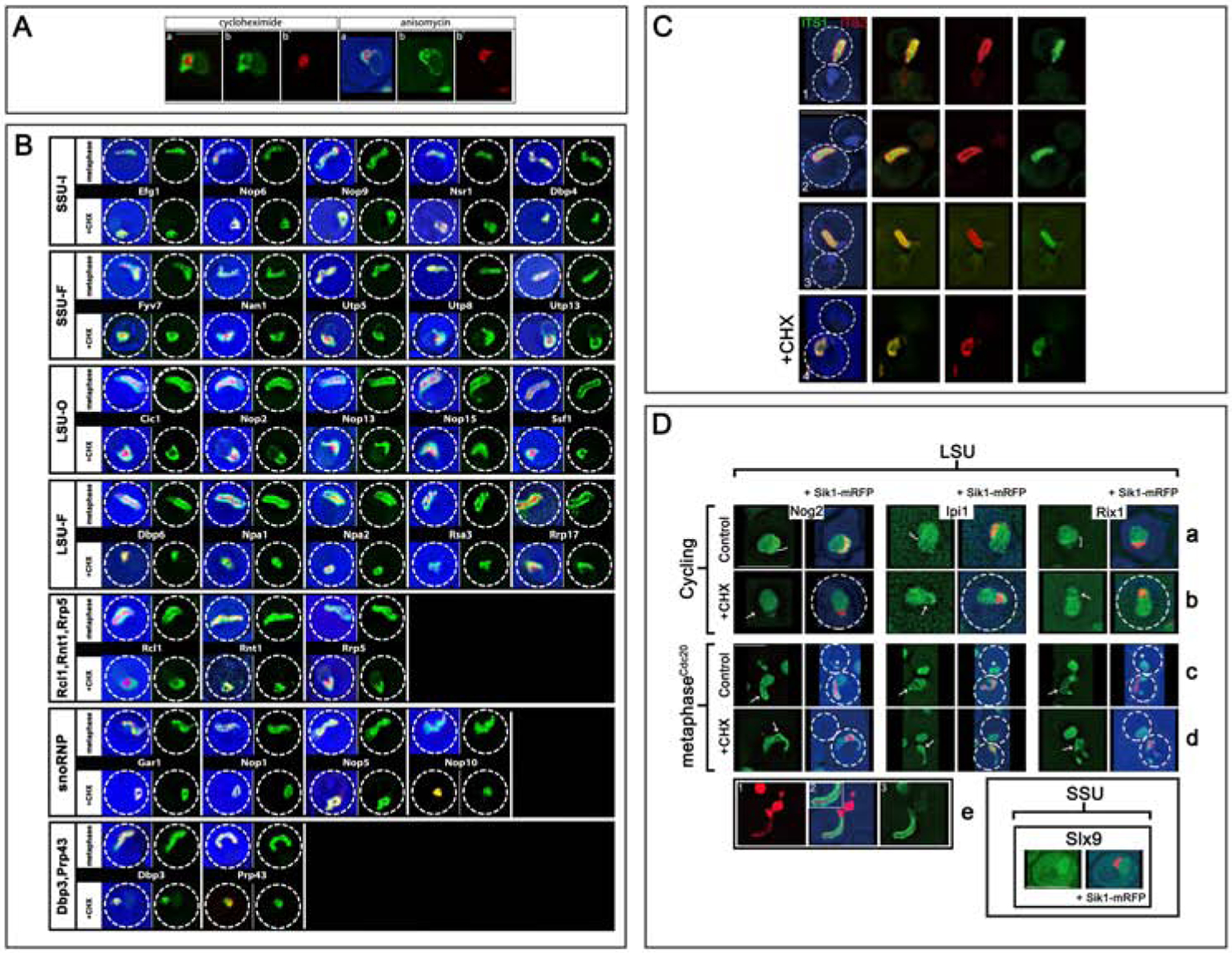
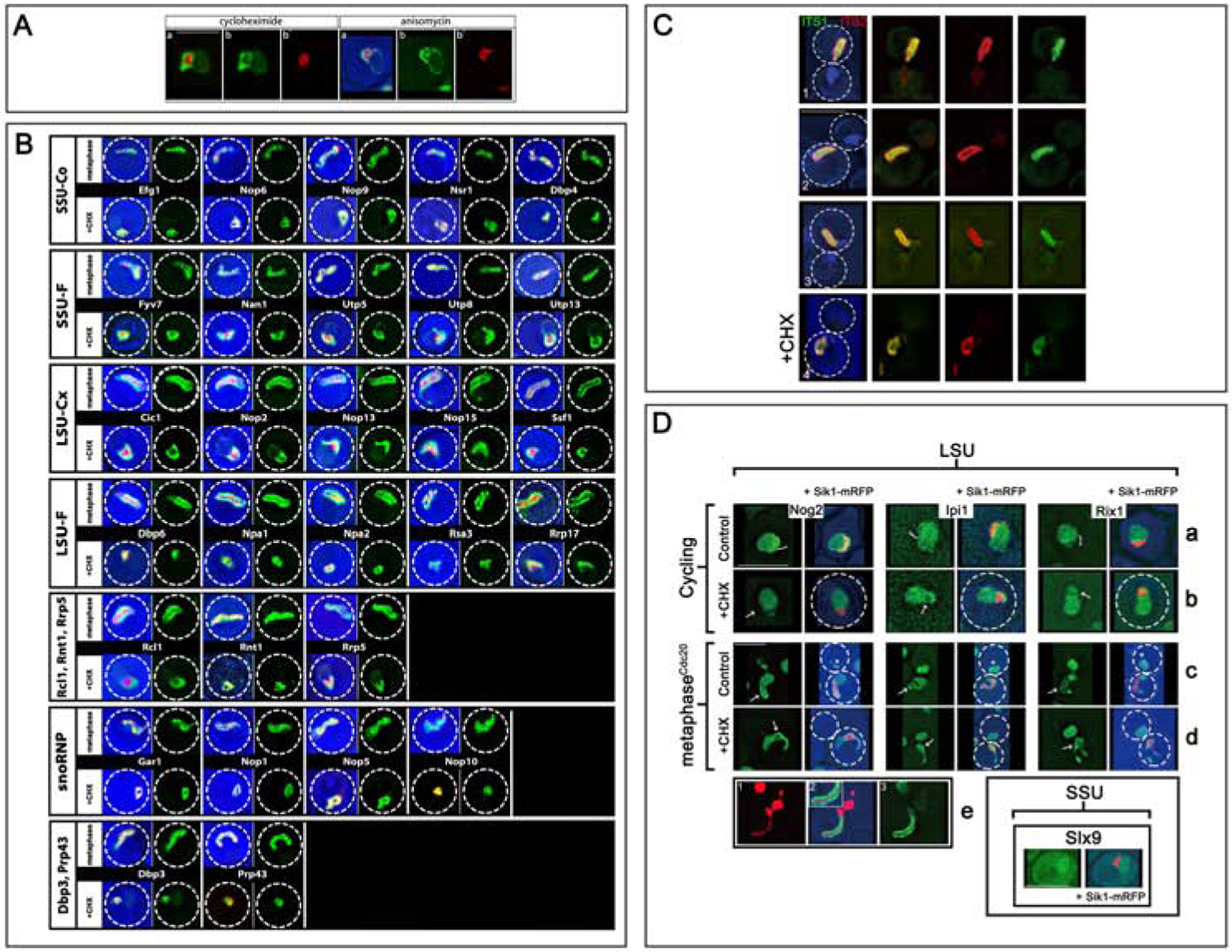
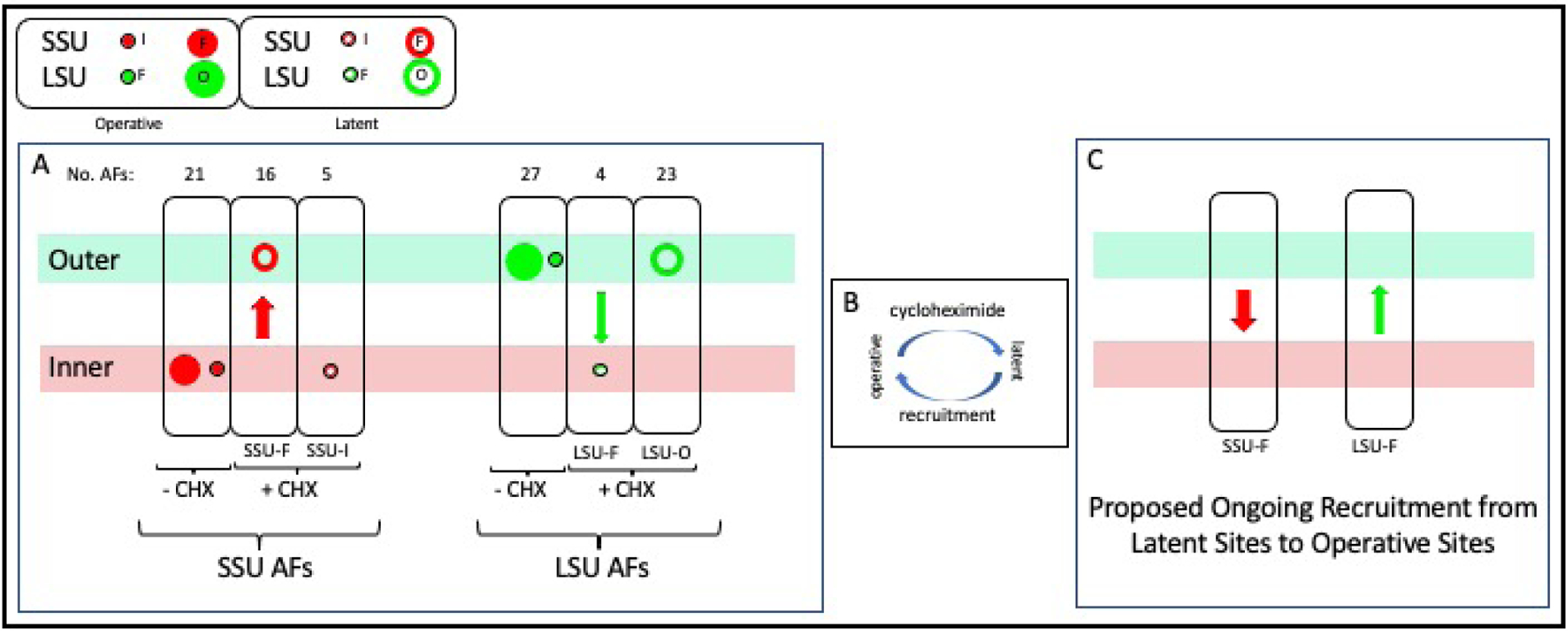
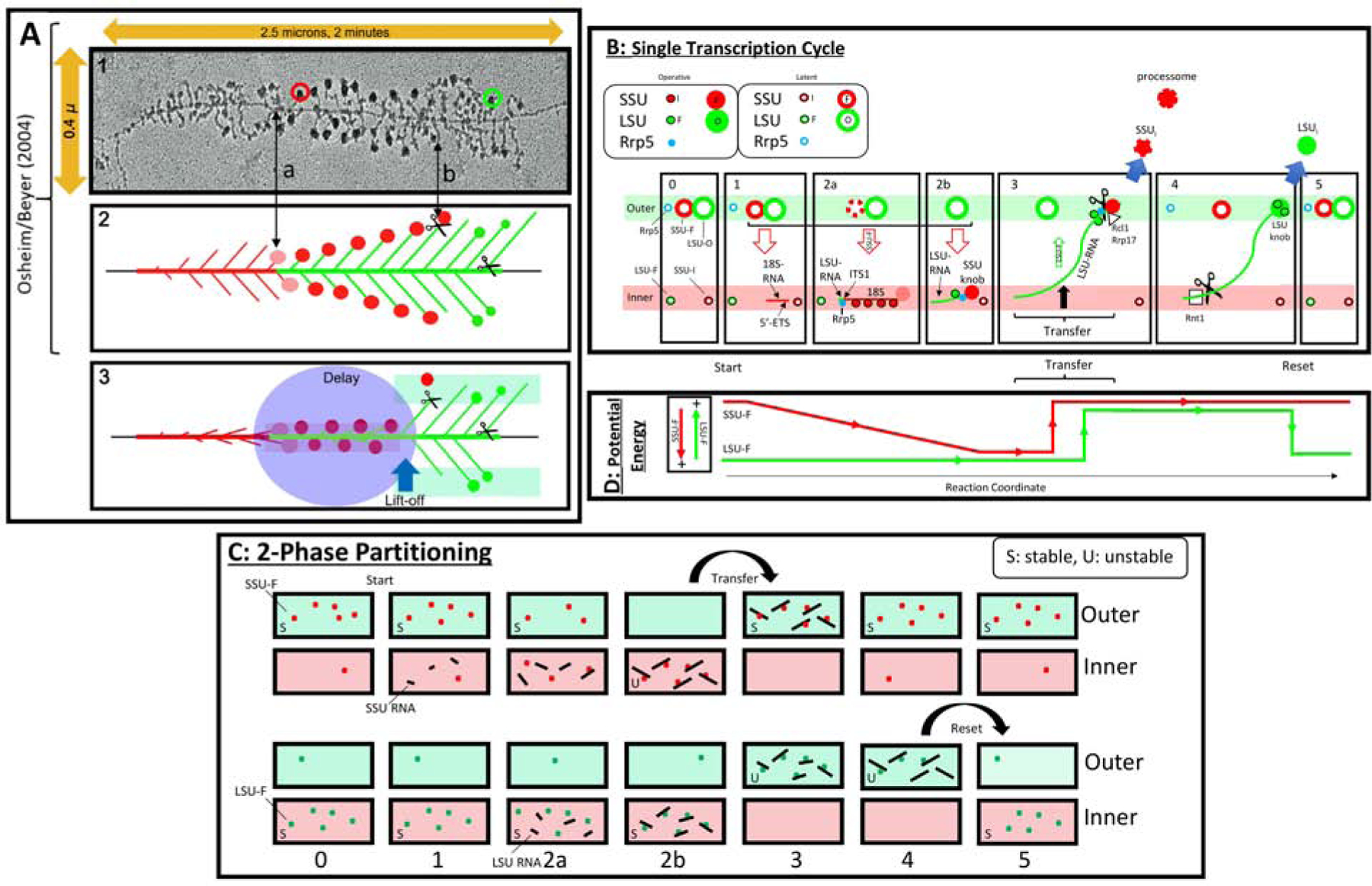
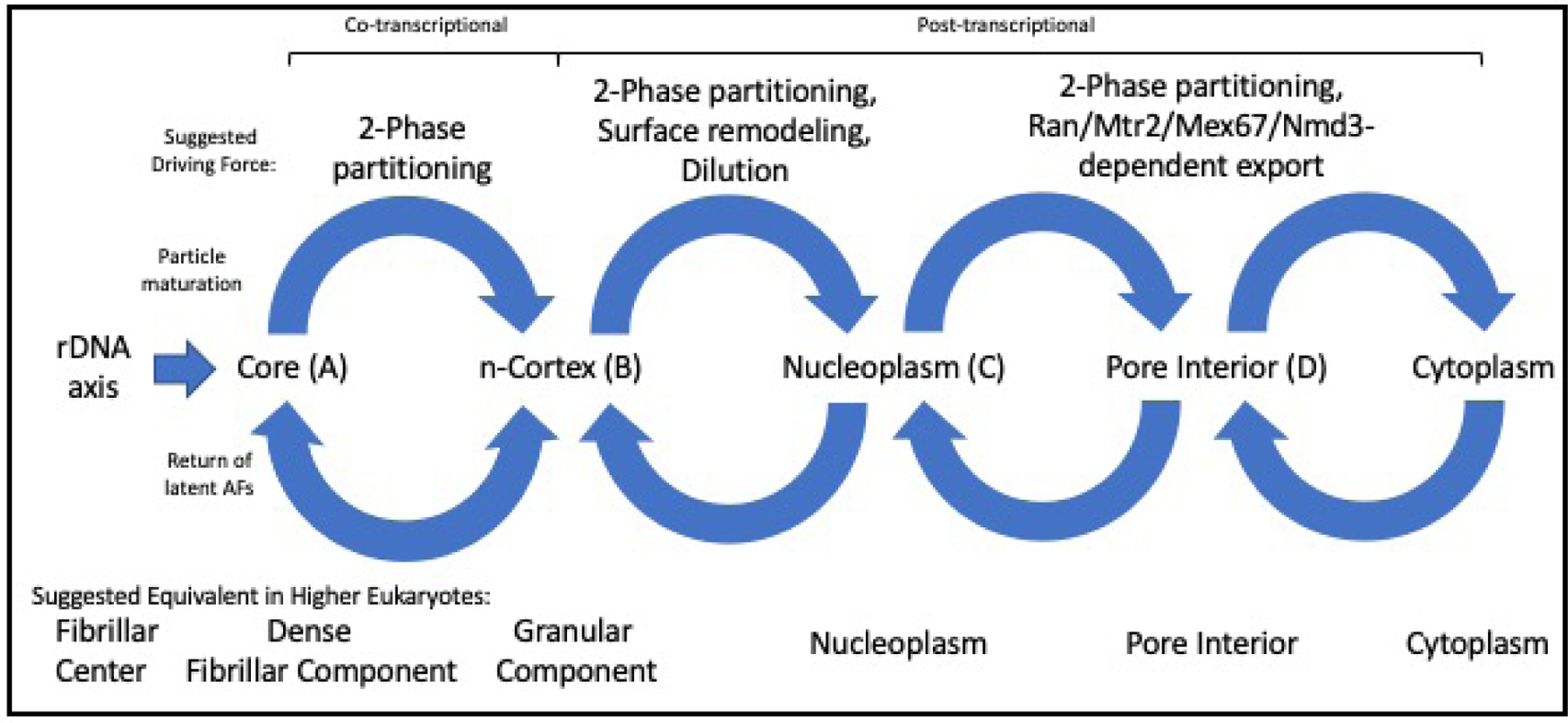
In ribosomal DNA (rDNA) repeats, sequences encoding small-subunit (SSU) rRNA precede those encoding large-subunit (LSU) rRNAs. Processing the composite transcript and subunit assembly requires >100 subunit-specific nucleolar assembly factors (AFs). To investigate the functional organization of the nucleolus, we localized AFs in S. cerevisiae in which the rDNA axis was "linearized" to reduce its dimensionality, thereby revealing its coaxial organization. In this situation, rRNA synthesis and processing continue. The axis is embedded in an inner layer/phase of SSU AFs that is surrounded by an outer layer/phase of LSU AFs. When subunit production is inhibited, subsets of AFs differentially relocate between the inner and outer layers, as expected if there is a cycle of repeated relocation whereby "latent" AFs become "operative" when recruited to nascent subunits. Recognition of AF cycling and localization of segments of rRNA make it possible to infer the existence of assembly intermediates that span between the inner and outer layers and to chart the cotranscriptional assembly of each subunit. AF cycling also can explain how having more than one protein phase in the nucleolus makes possible "vectorial 2-phase partitioning" as a driving force for relocation of nascent rRNPs. Because nucleoplasmic AFs are also present in the outer layer, we propose that critical surface remodeling occurs at this site, thereby partitioning subunit precursors into the nucleoplasm for post-transcriptional maturation. Comparison to observations on higher eukaryotes shows that the coaxial paradigm is likely to be applicable for the many other organisms that have rDNA repeats.
Keywords: 2-phase partitioning; assembly factors; nucleolar domain separation; nucleolar subcompartments; nucleolus; protein phase; rDNA; ribosomal subunit biogenesis; yeast.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Production of nascent ribosome precursors within the nucleolar microenvironment of Saccharomyces cerevisiae.Genetics. 2022 Jul 4;221(3):iyac070. doi: 10.1093/genetics/iyac070. Genetics. 2022. PMID: 35657327 Free PMC article.
-
Assembly and functional organization of the nucleolus: ultrastructural analysis of Saccharomyces cerevisiae mutants.Mol Biol Cell. 2000 Jun;11(6):2175-89. doi: 10.1091/mbc.11.6.2175. Mol Biol Cell. 2000. PMID: 10848637 Free PMC article.
-
How the concentric organization of the nucleolus and chromatin ensures accuracy of ribosome biogenesis and drives transport.Genetics. 2025 Apr 17;229(4):iyaf030. doi: 10.1093/genetics/iyaf030. Genetics. 2025. PMID: 40152466 Review.
-
Ribosomal DNA and the nucleolus at the heart of aging.Trends Biochem Sci. 2022 Apr;47(4):328-341. doi: 10.1016/j.tibs.2021.12.007. Epub 2022 Jan 18. Trends Biochem Sci. 2022. PMID: 35063340 Review.
-
Links between nucleolar activity, rDNA stability, aneuploidy and chronological aging in the yeast Saccharomyces cerevisiae.Biogerontology. 2014 Jun;15(3):289-316. doi: 10.1007/s10522-014-9499-y. Epub 2014 Apr 8. Biogerontology. 2014. PMID: 24711086 Free PMC article.
Cited by
-
Production of nascent ribosome precursors within the nucleolar microenvironment of Saccharomyces cerevisiae.Genetics. 2022 Jul 4;221(3):iyac070. doi: 10.1093/genetics/iyac070. Genetics. 2022. PMID: 35657327 Free PMC article.
-
Targeting of Hmo1 to subcompartments of the budding yeast nucleolus.Mol Biol Cell. 2023 Mar 1;34(3):ar22. doi: 10.1091/mbc.E22-07-0261. Epub 2023 Jan 25. Mol Biol Cell. 2023. PMID: 36696177 Free PMC article.
-
New insights into nuclear import and nucleolar localization of yeast RNA exosome subunits.Mol Biol Cell. 2025 Jun 1;36(6):ar69. doi: 10.1091/mbc.E25-02-0078. Epub 2025 Apr 23. Mol Biol Cell. 2025. PMID: 40266794
-
Cell cycle arrest of S. cerevisiae in conjunction with labeling of the cell wall.STAR Protoc. 2021 Jul 7;2(3):100646. doi: 10.1016/j.xpro.2021.100646. eCollection 2021 Sep 17. STAR Protoc. 2021. PMID: 34286287 Free PMC article.
-
The dual nature of the nucleolus.Genes Dev. 2022 Jul 1;36(13-14):765-769. doi: 10.1101/gad.349748.122. Genes Dev. 2022. PMID: 36342833 Free PMC article. Review.
References
-
- Nerurkar P, Altvater M, Gerhardy S, Schutz S, Fischer U, Weirich C, et al. Eukaryotic Ribosome Assembly and Nuclear Export. Int Rev Cell Mol Biol. 2015;319:107–40. - PubMed
-
- Pena C, Hurt E, Panse VG. Eukaryotic ribosome assembly, transport and quality control. Nature structural & molecular biology. 2017;24(9):689–99. - PubMed
-
- Bassler J, Hurt E. Eukaryotic Ribosome Assembly. Annu Rev Biochem. 2019;88:281–306. - PubMed
-
- Osheim YN, French SL, Keck KM, Champion EA, Spasov K, Dragon F, et al. Pre-18S ribosomal RNA is structurally compacted into the SSU processome prior to being cleaved from nascent transcripts in Saccharomyces cerevisiae. Molecular cell. 2004;16(6):943–54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
