

Characterization of CRN-Like Genes From Plasmopara viticola: Searching for the Most Virulent Ones
- PMID: 33868192
- PMCID: PMC8044898
- DOI: 10.3389/fmicb.2021.632047
Characterization of CRN-Like Genes From Plasmopara viticola: Searching for the Most Virulent Ones
Abstract
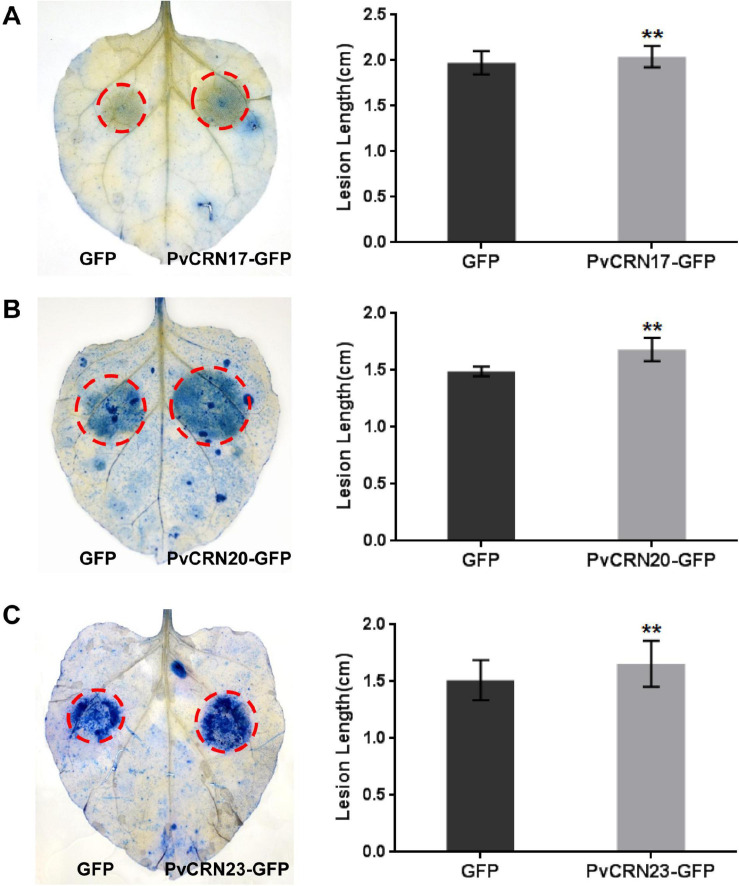
Grapevine downy mildew is an insurmountable disease that endangers grapevine production and the wine industry worldwide. The causal agent of the disease is the obligate biotrophic oomycete Plasmopara viticola, for which the pathogenic mechanism remains largely unknown. Crinkling and necrosis proteins (CRN) are an ancient class of effectors utilized by pathogens, including oomycetes, that interfere with host plant defense reactions. In this study, 27 CRN-like genes were cloned from the P. viticola isolate YL genome, hereafter referred to as PvCRN genes, and characterized in silico and in planta. PvCRN genes in 'YL' share high sequence identities with their ortholog genes in the other three previously sequenced P. viticola isolates. Sequence divergence among the genes in the PvCRN family indicates that different PvCRN genes have different roles. Phylogenetic analysis of the PvCRN and the CRN proteins encoded by genes in the P. halstedii genome suggests that various functions might have been acquired by the CRN superfamily through independent evolution of Plasmopara species. When transiently expressed in plant cells, the PvCRN protein family shows multiple subcellular localizations. None of the cloned PvCRN proteins induced hypersensitive response (HR)-like cell death on the downy mildew-resistant grapevine Vitis riparia. This was in accordance with the result that most PvCRN proteins, except PvCRN11, failed to induce necrosis in Nicotiana benthamiana. Pattern-triggered immunity (PTI) induced by INF1 was hampered by several PvCRN proteins. In addition, 15 PvCRN proteins prevented Bax-induced plant programmed cell death. Among the cell death-suppressing members, PvCRN17, PvCRN20, and PvCRN23 were found to promote the susceptibility of N. benthamiana to Phytophthora capsici, which is a semi-biotrophic oomycete. Moreover, the nucleus-targeting member, PvCRN19, promoted the susceptibility of N. benthamiana to P. capsici. Therefore, these PvCRN proteins were estimated to be virulent effectors involved in the pathogenicity of P. viticola YL. Collectively, this study provides comprehensive insight into the CRN effector repertoire of P. viticola YL, which will help further elucidate the molecular mechanisms of the pathogenesis of grapevine downy mildew.
Keywords: CRN effectors; Plasmopara viticola; Vitis; cell death; virulence.
Copyright © 2021 Xiang, Yin, Niu, Chen, Liu, Shang, Fu, Liu, Ma and Xu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures









Similar articles
-
The cytosolic iron-sulphur cluster assembly mechanism in grapevine is one target of a virulent Crinkler effector from Plasmopara viticola.Mol Plant Pathol. 2022 Dec;23(12):1792-1806. doi: 10.1111/mpp.13266. Epub 2022 Sep 7. Mol Plant Pathol. 2022. PMID: 36071584 Free PMC article.
-
The pathogenicity of Plasmopara viticola: a review of evolutionary dynamics, infection strategies and effector molecules.BMC Plant Biol. 2024 Apr 24;24(1):327. doi: 10.1186/s12870-024-05037-0. BMC Plant Biol. 2024. PMID: 38658826 Free PMC article. Review.
-
Plasmopara viticola effector PvCRN11 induces disease resistance to downy mildew in grapevine.Plant J. 2024 Feb;117(3):873-891. doi: 10.1111/tpj.16534. Epub 2023 Nov 11. Plant J. 2024. PMID: 37950600
-
ADP-ribosylation factor 1 (ARF1) protein interacts with elicitor PvNLP7 from Plasmopara viticola to mediate PvNLP7-triggered immunity.Plant Sci. 2024 Oct;347:112194. doi: 10.1016/j.plantsci.2024.112194. Epub 2024 Jul 14. Plant Sci. 2024. PMID: 39009307
-
Advances in understanding grapevine downy mildew: From pathogen infection to disease management.Mol Plant Pathol. 2024 Jan;25(1):e13401. doi: 10.1111/mpp.13401. Epub 2023 Nov 22. Mol Plant Pathol. 2024. PMID: 37991155 Free PMC article. Review.
Cited by
-
The cytosolic iron-sulphur cluster assembly mechanism in grapevine is one target of a virulent Crinkler effector from Plasmopara viticola.Mol Plant Pathol. 2022 Dec;23(12):1792-1806. doi: 10.1111/mpp.13266. Epub 2022 Sep 7. Mol Plant Pathol. 2022. PMID: 36071584 Free PMC article.
-
Differing Responses to Phytophthora cinnamomi Infection in Susceptible and Partially Resistant Persea americana (Mill.) Rootstocks: A Case for the Role of Receptor-Like Kinases and Apoplastic Proteases.Front Plant Sci. 2022 Jun 28;13:928176. doi: 10.3389/fpls.2022.928176. eCollection 2022. Front Plant Sci. 2022. PMID: 35837458 Free PMC article.
-
Functional Analysis of KPvRxLR27, a Novel Plasmopara viticola Effector from a Korean Isolate, and Its Role in Hypersensitive Response.Plant Pathol J. 2025 Feb;41(1):28-37. doi: 10.5423/PPJ.OA.09.2024.0141. Epub 2024 Dec 16. Plant Pathol J. 2025. PMID: 39676500 Free PMC article.
-
The pathogenicity of Plasmopara viticola: a review of evolutionary dynamics, infection strategies and effector molecules.BMC Plant Biol. 2024 Apr 24;24(1):327. doi: 10.1186/s12870-024-05037-0. BMC Plant Biol. 2024. PMID: 38658826 Free PMC article. Review.
-
Xylanase VmXyl2 is involved in the pathogenicity of Valsa mali by regulating xylanase activity and inducing cell necrosis.Front Plant Sci. 2024 Apr 29;15:1342714. doi: 10.3389/fpls.2024.1342714. eCollection 2024. Front Plant Sci. 2024. PMID: 38745923 Free PMC article.
References
-
- Armstrong M. R., Whisson S. C., Pritchard L., Bos J. I., Venter E., Avrova A. O., et al. (2005). An ancestral oomycete locus contains late blight avirulence gene Avr3a, encoding a protein that is recognized in the host cytoplasm. Proc. Natl. Acad. Sci. U.S.A. 21 7766–7771. 10.1073/pnas.0500113102 - DOI - PMC - PubMed
-
- Bailey K., Cevik V., Holton N., Byrne-Richardson J., Sohn K. H., Coates M., et al. (2011). Molecular cloning of ATR5(Emoy2) from Hyaloperonospora arabidopsidis, an avirulence determinant that triggers RPP5-mediated defense in Arabidopsis. Mol. Plant Microbe Interact. 7 827–838. 10.1094/MPMI-12-10-0278 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous