

Telomere Replication: Solving Multiple End Replication Problems
- PMID: 33869233
- PMCID: PMC8047117
- DOI: 10.3389/fcell.2021.668171
Telomere Replication: Solving Multiple End Replication Problems
Abstract
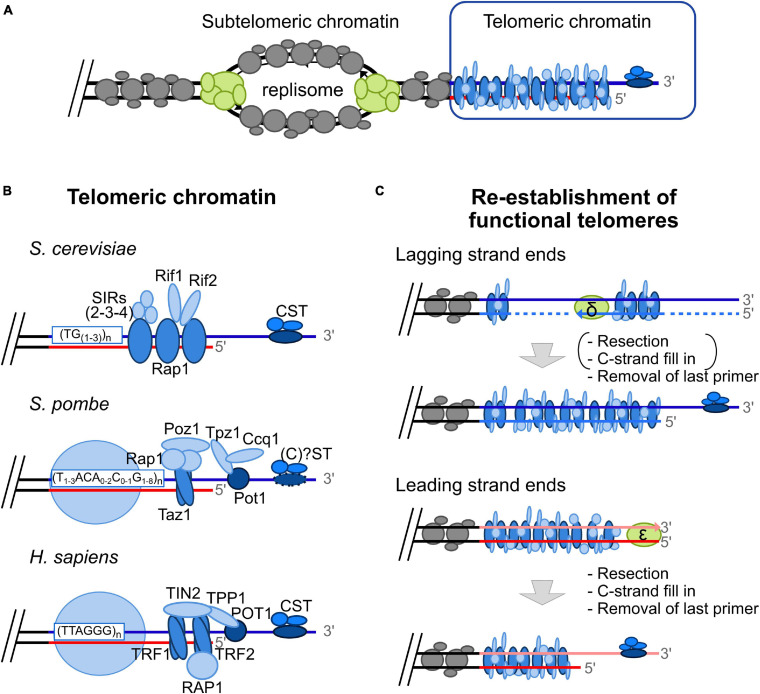
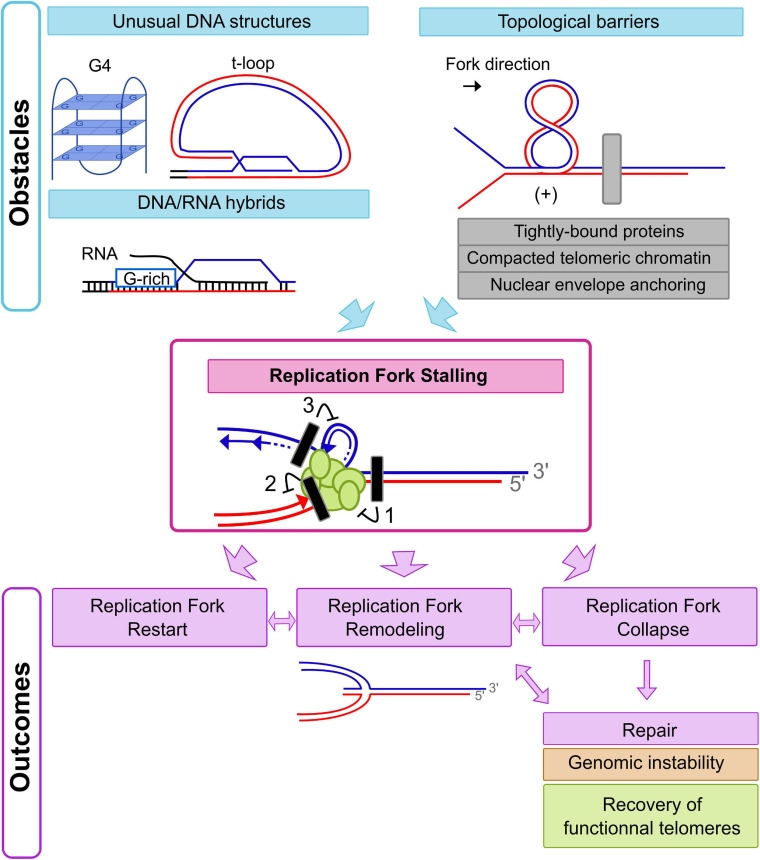
Eukaryotic genomes are highly complex and divided into linear chromosomes that require end protection from unwarranted fusions, recombination, and degradation in order to maintain genomic stability. This is accomplished through the conserved specialized nucleoprotein structure of telomeres. Due to the repetitive nature of telomeric DNA, and the unusual terminal structure, namely a protruding single stranded 3' DNA end, completing telomeric DNA replication in a timely and efficient manner is a challenge. For example, the end replication problem causes a progressive shortening of telomeric DNA at each round of DNA replication, thus telomeres eventually lose their protective capacity. This phenomenon is counteracted by the recruitment and the activation at telomeres of the specialized reverse transcriptase telomerase. Despite the importance of telomerase in providing a mechanism for complete replication of telomeric ends, the majority of telomere replication is in fact carried out by the conventional DNA replication machinery. There is significant evidence demonstrating that progression of replication forks is hampered at chromosomal ends due to telomeric sequences prone to form secondary structures, tightly DNA-bound proteins, and the heterochromatic nature of telomeres. The telomeric loop (t-loop) formed by invasion of the 3'-end into telomeric duplex sequences may also impede the passage of replication fork. Replication fork stalling can lead to fork collapse and DNA breaks, a major cause of genomic instability triggered notably by unwanted repair events. Moreover, at chromosomal ends, unreplicated DNA distal to a stalled fork cannot be rescued by a fork coming from the opposite direction. This highlights the importance of the multiple mechanisms involved in overcoming fork progression obstacles at telomeres. Consequently, numerous factors participate in efficient telomeric DNA duplication by preventing replication fork stalling or promoting the restart of a stalled replication fork at telomeres. In this review, we will discuss difficulties associated with the passage of the replication fork through telomeres in both fission and budding yeasts as well as mammals, highlighting conserved mechanisms implicated in maintaining telomere integrity during replication, thus preserving a stable genome.
Keywords: DNA replication; genome stability; replication fork stability; telomeres; telomeric chromatin.
Copyright © 2021 Bonnell, Pasquier and Wellinger.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources