

Time of the day prioritizes the pool of translating mRNAs in response to heat stress
- PMID: 33871647
- PMCID: PMC8364243
- DOI: 10.1093/plcell/koab113
Time of the day prioritizes the pool of translating mRNAs in response to heat stress
Abstract
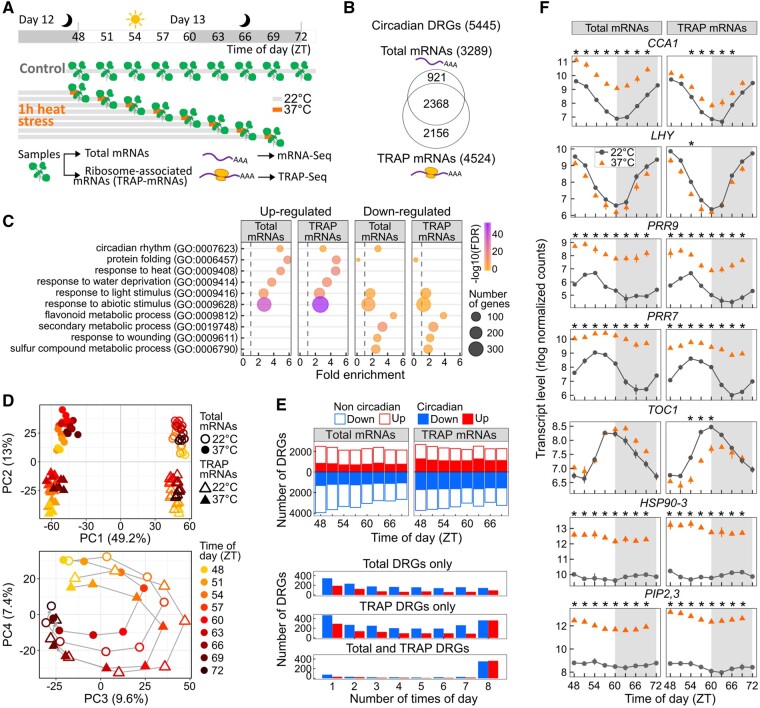
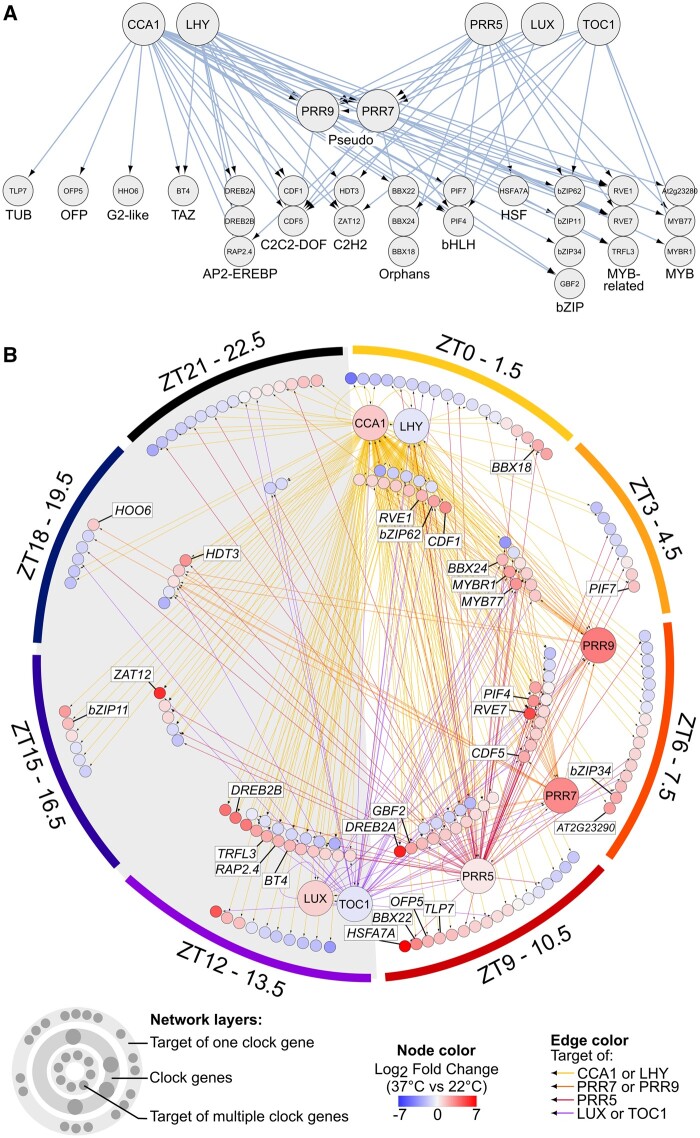
The circadian clock helps organisms to anticipate and coordinate gene regulatory responses to changes in environmental stimuli. Under growth limiting temperatures, the time of the day modulates the accumulation of polyadenylated mRNAs. In response to heat stress, plants will conserve energy and selectively translate mRNAs. How the clock and/or the time of the day regulates polyadenylated mRNAs bound by ribosomes in response to heat stress is unknown. In-depth analysis of Arabidopsis thaliana translating mRNAs found that the time of the day gates the response of approximately one-third of the circadian-regulated heat-responsive translatome. Specifically, the time of the day and heat stress interact to prioritize the pool of mRNAs in cue to be translated. For a subset of mRNAs, we observed a stronger gated response during the day, and preferentially before the peak of expression. We propose previously overlooked transcription factors (TFs) as regulatory nodes and show that the clock plays a role in the temperature response for select TFs. When the stress was removed, the redefined priorities for translation recovered within 1 h, though slower recovery was observed for abiotic stress regulators. Through hierarchical network connections between clock genes and prioritized TFs, our work provides a framework to target key nodes underlying heat stress tolerance throughout the day.
© American Society of Plant Biologists 2021. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures







References
-
- Adams S, Grundy J, Veflingstad SR, Dyer NP, Hannah MA, Ott S, Carré IA (2018) Circadian control of abscisic acid biosynthesis and signalling pathways revealed by genome-wide analysis of LHY binding targets. New Phytol 220:893–907 - PubMed
-
- Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Ser B 57:289–300
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
