

Bacterial persisters are a stochastically formed subpopulation of low-energy cells
- PMID: 33872303
- PMCID: PMC8084331
- DOI: 10.1371/journal.pbio.3001194
Bacterial persisters are a stochastically formed subpopulation of low-energy cells
Abstract
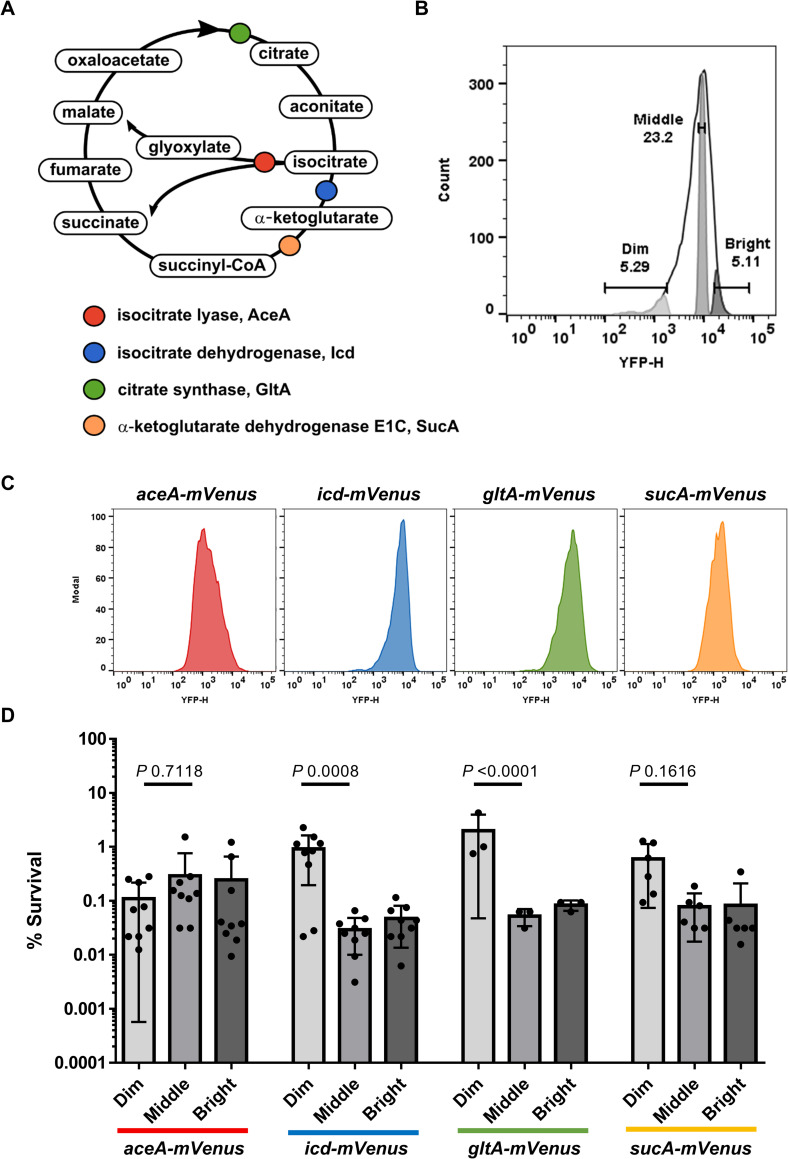
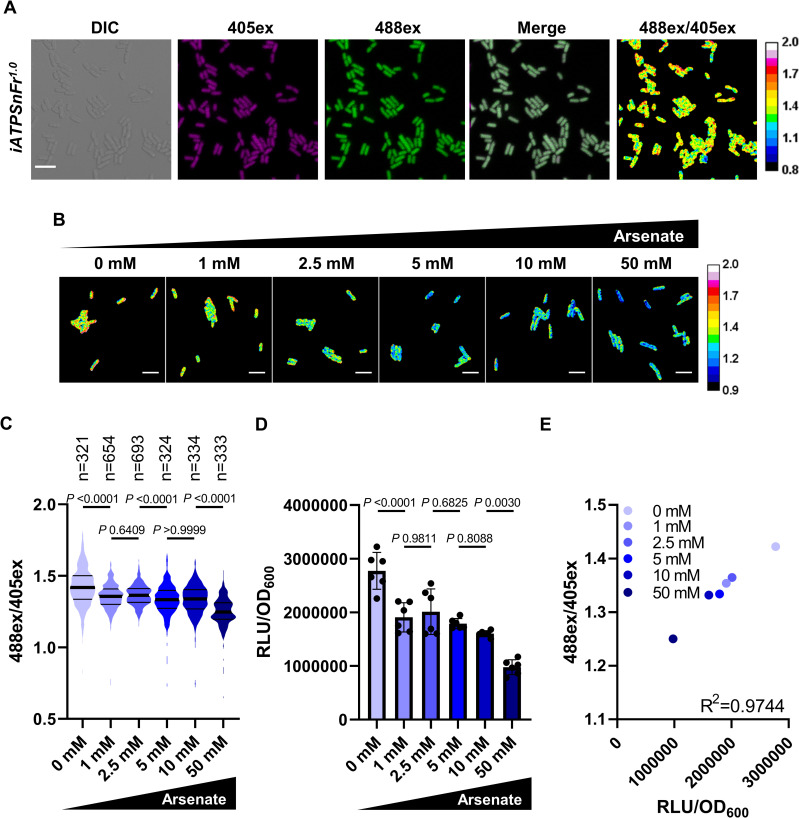
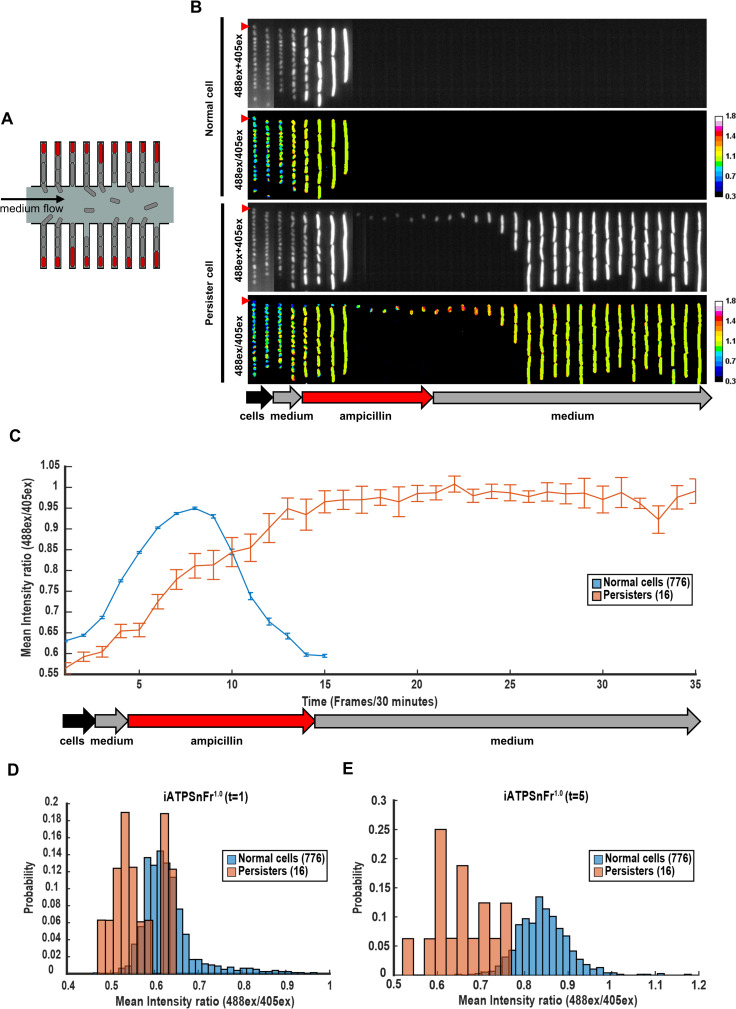
Persisters represent a small subpopulation of non- or slow-growing bacterial cells that are tolerant to killing by antibiotics. Despite their prominent role in the recalcitrance of chronic infections to antibiotic therapy, the mechanism of their formation has remained elusive. We show that sorted cells of Escherichia coli with low levels of energy-generating enzymes are better able to survive antibiotic killing. Using microfluidics time-lapse microscopy and a fluorescent reporter for in vivo ATP measurements, we find that a subpopulation of cells with a low level of ATP survives killing by ampicillin. We propose that these low ATP cells are formed stochastically as a result of fluctuations in the abundance of energy-generating components. These findings point to a general "low energy" mechanism of persister formation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Conlon BP, Rowe SE, Gandt AB, Nuxoll AS, Donegan NP, Zalis EA, et al. Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat Microbiol. 2016:1. https://www.nature.com/articles/nmicrobiol201651 - PMC - PubMed
-
- Goormaghtigh F, Fraikin N, Putrins M, Hallaert T, Hauryliuk V, Garcia-Pino A, et al. Reassessing the role of type II toxin-antitoxin systems in formation of Escherichia coli type II persister ells. MBio. 2018;9(3). https://mbio.asm.org/content/9/3/e00640-18.long - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
