

Implication of repeat insertion domains in the trans-activity of the long non-coding RNA ANRIL
- PMID: 33872355
- PMCID: PMC8136789
- DOI: 10.1093/nar/gkab245
Implication of repeat insertion domains in the trans-activity of the long non-coding RNA ANRIL
Abstract
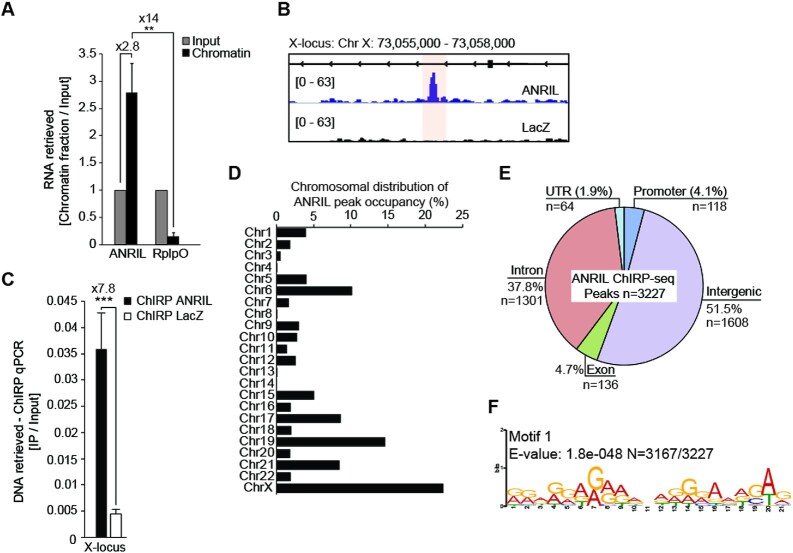
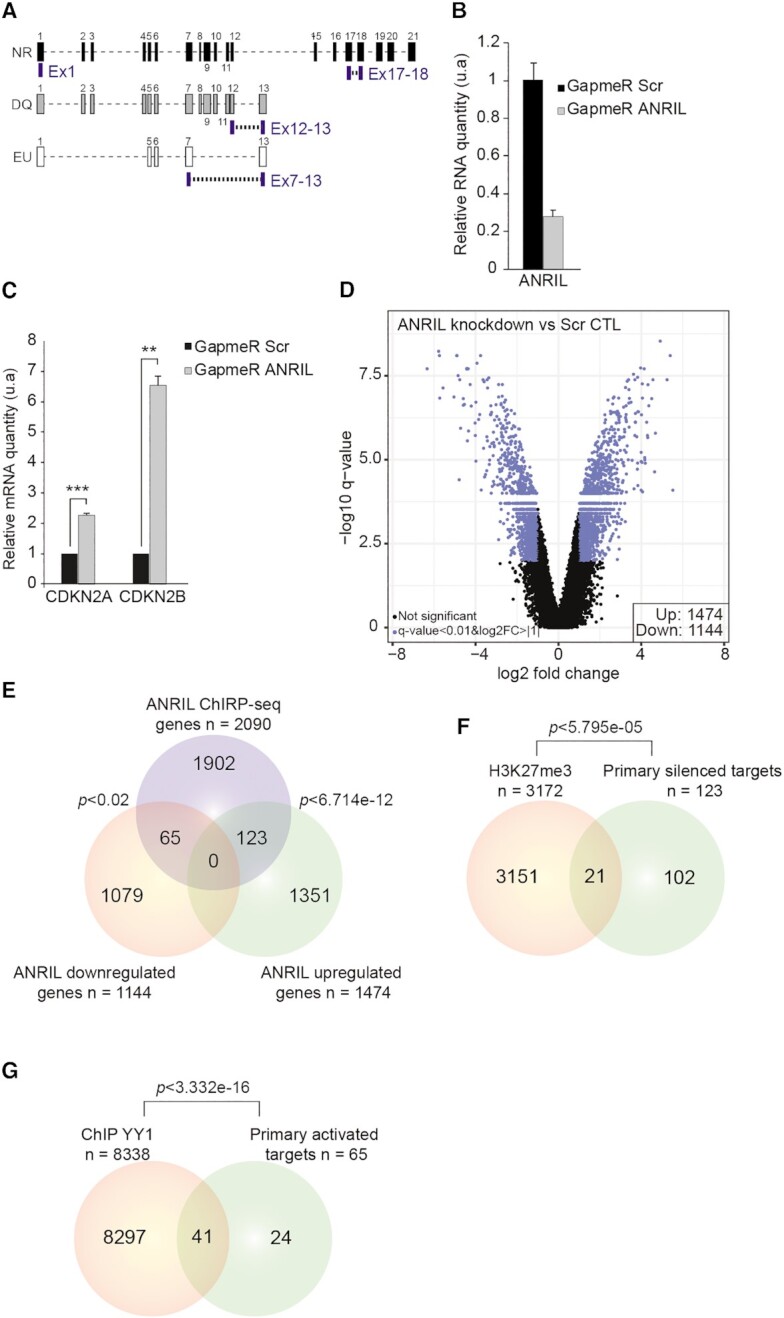
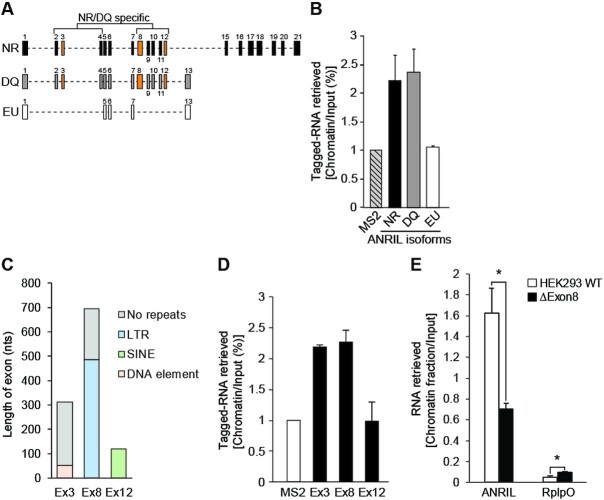
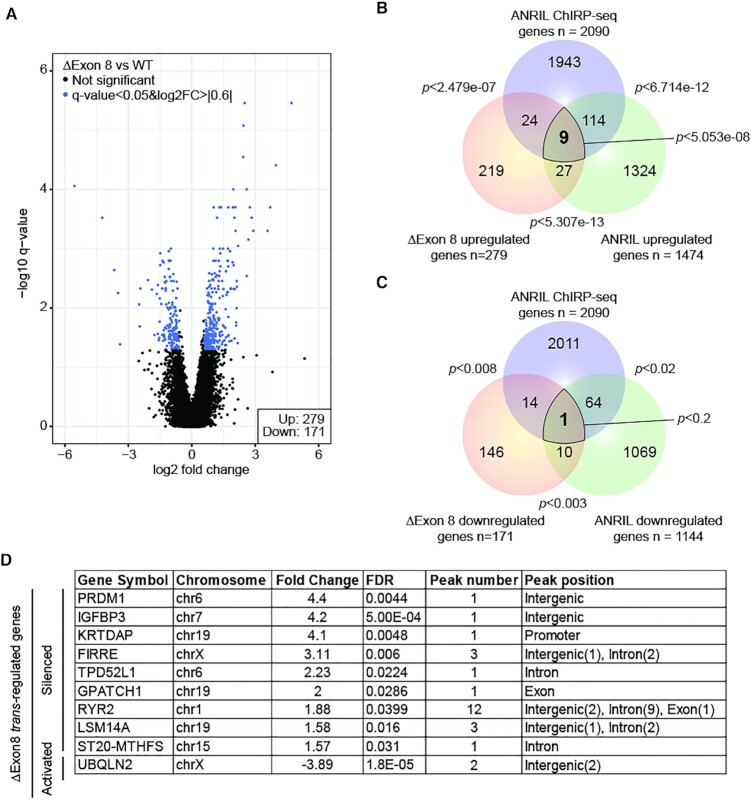
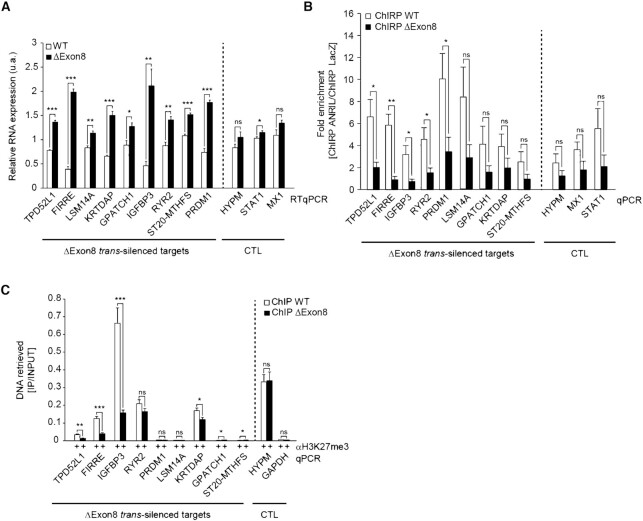
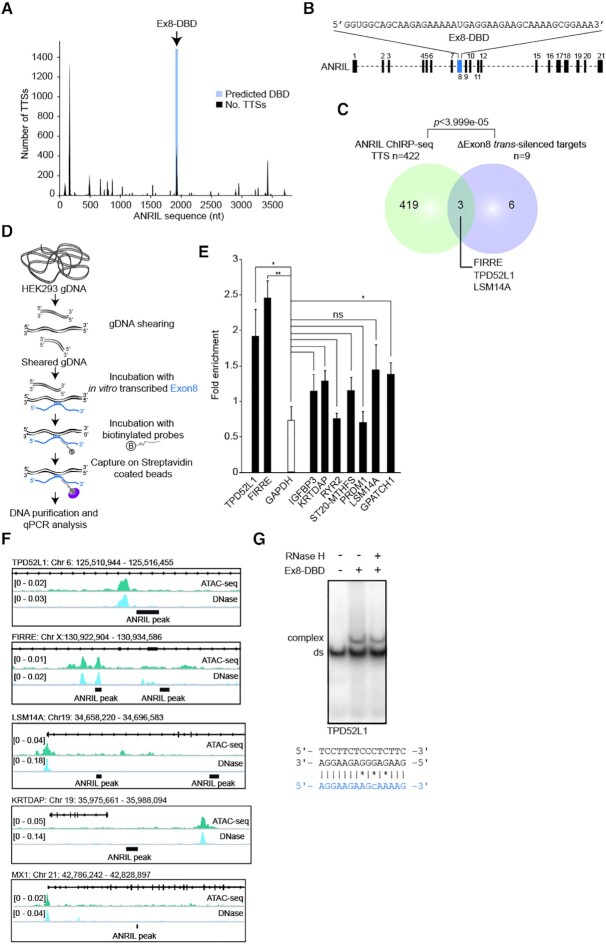
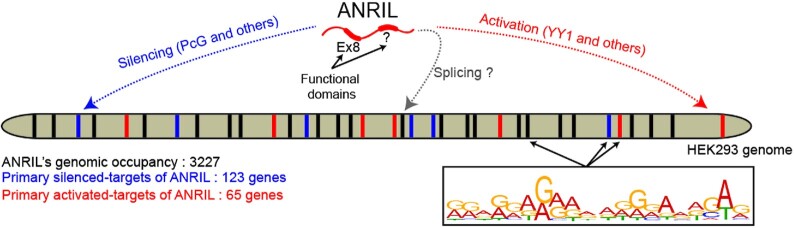
Long non-coding RNAs have emerged as critical regulators of cell homeostasis by modulating gene expression at chromatin level for instance. Here, we report that the lncRNA ANRIL, associated with several pathologies, binds to thousands of loci dispersed throughout the mammalian genome sharing a 21-bp motif enriched in G/A residues. By combining ANRIL genomic occupancy with transcriptomic analysis, we established a list of 65 and 123 genes potentially directly activated and silenced by ANRIL in trans, respectively. We also found that Exon8 of ANRIL, mainly made of transposable elements, contributes to ANRIL genomic association and consequently to its trans-activity. Furthermore, we showed that Exon8 favors ANRIL's association with the FIRRE, TPD52L1 and IGFBP3 loci to modulate their expression through H3K27me3 deposition. We also investigated the mechanisms engaged by Exon8 to favor ANRIL's association with the genome. Our data refine ANRIL's trans-activity and highlight the functional importance of TEs on ANRIL's activity.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Mercer T.R., Dinger M.E., Mattick J.S.. Long non-coding RNAs: insights into functions. Nat. Rev. Genet. 2009; 10:155–159. - PubMed
-
- Wapinski O., Chang H.Y.. Long noncoding RNAs and human disease. Trends Cell Biol. 2011; 21:354–361. - PubMed
-
- Yao R.-W., Wang Y., Chen L.-L.. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019; 21:542–551. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
