

Mutations in Ehrlichia chaffeensis Genes ECH_0660 and ECH_0665 Cause Transcriptional Changes in Response to Zinc or Iron Limitation
- PMID: 33875547
- PMCID: PMC8316085
- DOI: 10.1128/JB.00027-21
Mutations in Ehrlichia chaffeensis Genes ECH_0660 and ECH_0665 Cause Transcriptional Changes in Response to Zinc or Iron Limitation
Abstract
Ehrlichia chaffeensis causes human monocytic ehrlichiosis by replicating within phagosomes of monocytes/macrophages. A function disruption mutation within the pathogen's ECH_0660 gene, which encodes a phage head-to-tail connector protein, resulted in the rapid clearance of the pathogen in vivo, while aiding in induction of sufficient immunity in a host to protect against wild-type infection challenge. In this study, we describe the characterization of a cluster of seven genes spanning from ECH_0659 to ECH_0665, which contained four genes encoding bacterial phage proteins, including the ECH_0660 gene. Assessment of the promoter region upstream of the first gene of the seven genes (ECH_0659) in Escherichia coli demonstrated transcriptional enhancement under zinc and iron starvation conditions. Furthermore, transcription of the seven genes was significantly higher under zinc and iron starvation conditions for E. chaffeensis carrying a mutation in the ECH_0660 gene compared to the wild-type pathogen. In contrast, for the ECH_0665 gene mutant with the function disruption, transcription from the genes was mostly similar to that of the wild type or was moderately downregulated. Recently, we reported that this mutation caused a minimal impact on the pathogen's in vivo growth, as it persisted similarly to the wild type. The current study is the first to describe how zinc and iron contribute to E. chaffeensis biology. Specifically, we demonstrated that the functional disruption in the gene encoding the phage head-to-tail connector protein in E. chaffeensis results in the enhanced transcription of seven genes, including those encoding phage proteins, under zinc and iron limitation. IMPORTANCE Ehrlichia chaffeensis, a tick-transmitted bacterium, causes human monocytic ehrlichiosis by replicating within phagosomes of monocytes/macrophages. A function disruption mutation within the pathogen's gene encoding a phage head-to-tail connector protein resulted in the rapid clearance of the pathogen in vivo, while aiding in induction of sufficient immunity in a host to protect against wild-type infection challenge. In the current study, we investigated if the functional disruption in the phage head-to-tail connector protein gene caused transcriptional changes resulting from metal ion limitations. This is the first study describing how zinc and iron may contribute to E. chaffeensis replication.
Keywords: Anaplasmataceae pathogens; Ehrlichia chaffeensis; Rickettsiales; iron; metal ion deficiency; metal ions; tick-borne diseases; zinc.
Figures

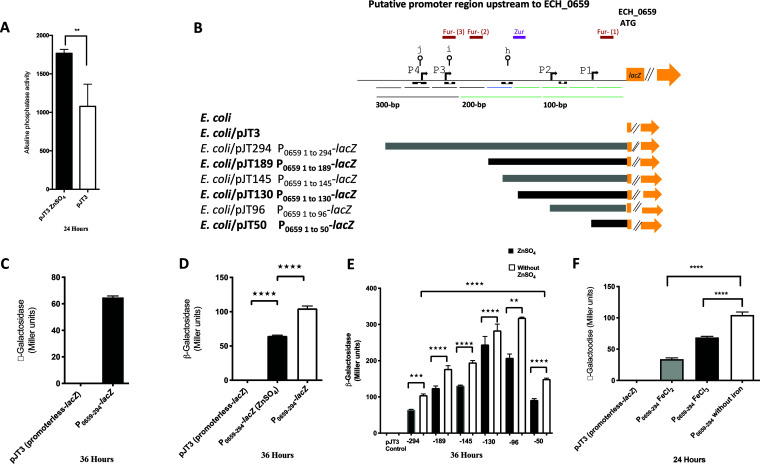
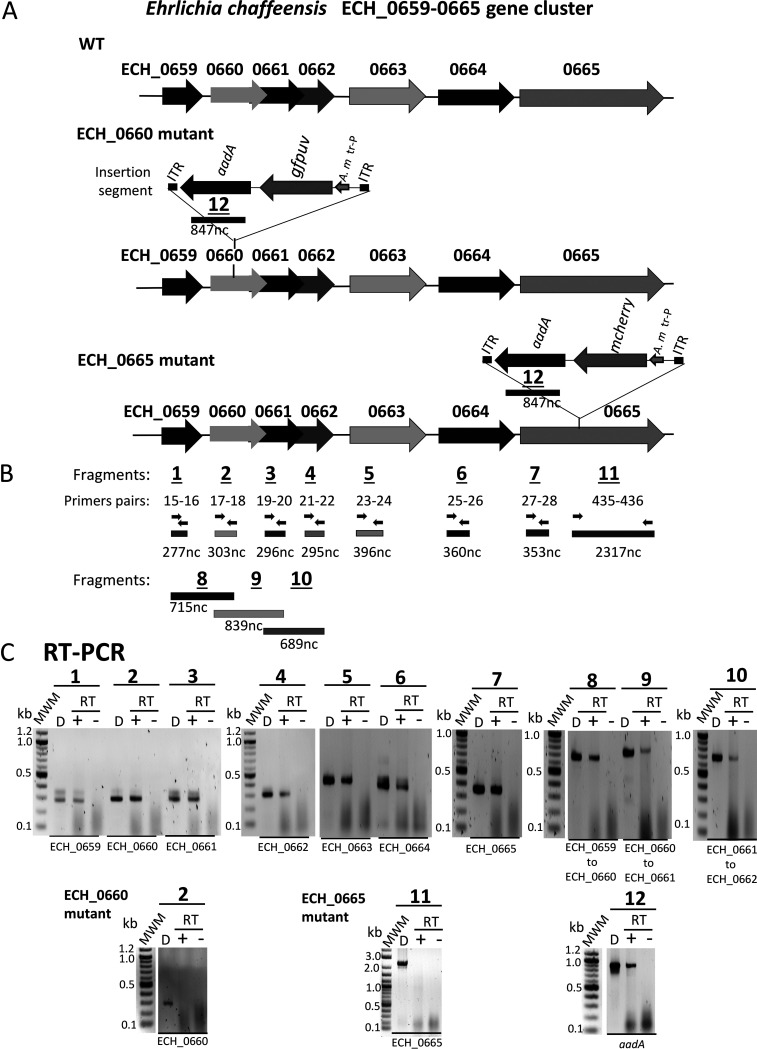
References
-
- Ewing SA, Dawson JE, Kocan AA, Barker RW, Warner CK, Panciera RJ, Fox JC, Kocan KM, Blouin EF. 1995. Experimental transmission of Ehrlichia chaffeensis (Rickettsiales: Ehrlichieae) among white-tailed deer by Amblyomma americanum (Acari: Ixodidae). J Med Entomol 32:368–374. 10.1093/jmedent/32.3.368. - DOI - PubMed
-
- Dumler JS, Barbet AF, Bekker CP, Dasch GA, Palmer GH, Ray SC, Rikihisa Y, Rurangirwa FR. 2001. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int J Syst Evol Microbiol 51:2145–2165. 10.1099/00207713-51-6-2145. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
