

Transcriptional profiling reveals signatures of latent developmental potential in Arabidopsis stomatal lineage ground cells
- PMID: 33875598
- PMCID: PMC8092560
- DOI: 10.1073/pnas.2021682118
Transcriptional profiling reveals signatures of latent developmental potential in Arabidopsis stomatal lineage ground cells
Abstract
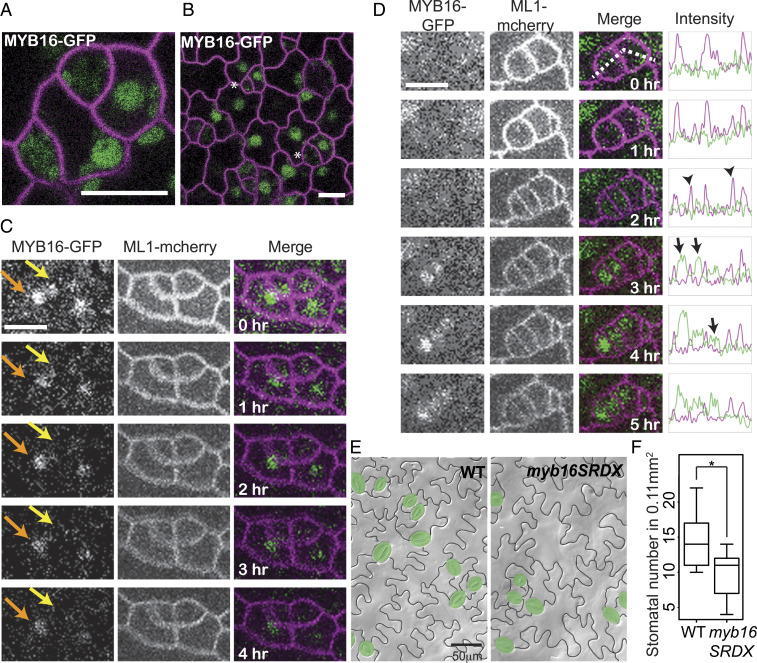
In many developmental contexts, cell lineages have variable or flexible potency to self-renew. What drives a cell to exit from a proliferative state and begin differentiation, or to retain the capacity to divide days or years later is not clear. Here we exploit the mixed potential of the stomatal lineage ground cell (SLGC) in the Arabidopsis leaf epidermis as a model to explore how cells might balance potential to differentiate with a reentry into proliferation. By generating transcriptomes of fluorescence-activated cell sorting-isolated populations that combinatorically define SLGCs and integrating these data with other stomatal lineage datasets, we find that SLGCs appear poised between proliferation and endoreduplication. Furthermore, we found the RNA polymerase II-related mediator complex interactor DEK and the transcription factor MYB16 accumulate differentially in the stomatal lineage and influence the extent of cell proliferation during leaf development. These findings suggest that SLGC latent potential is maintained by poising of the cell cycle machinery, as well as general and site-specific gene-expression regulators.
Keywords: Arabidopsis; MYB16 endoreduplication; stomatal lineage ground cell; transcriptional profile.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
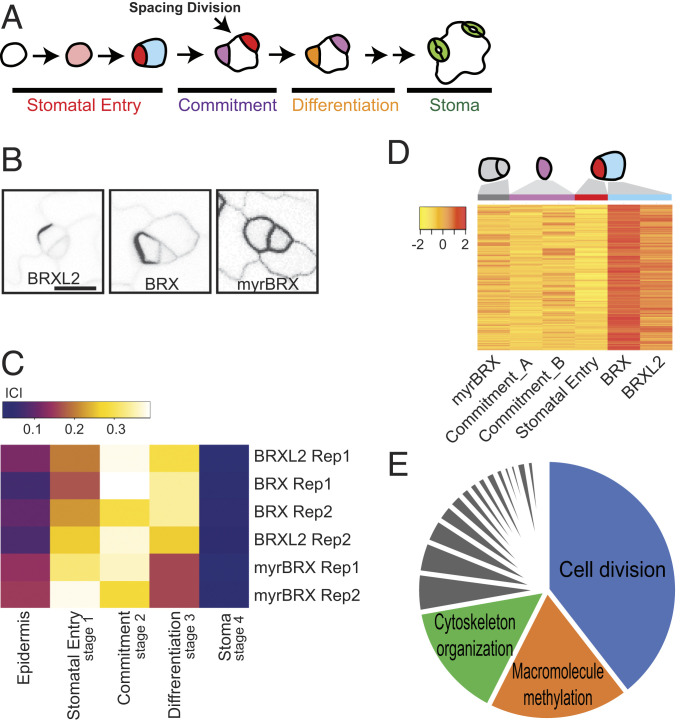
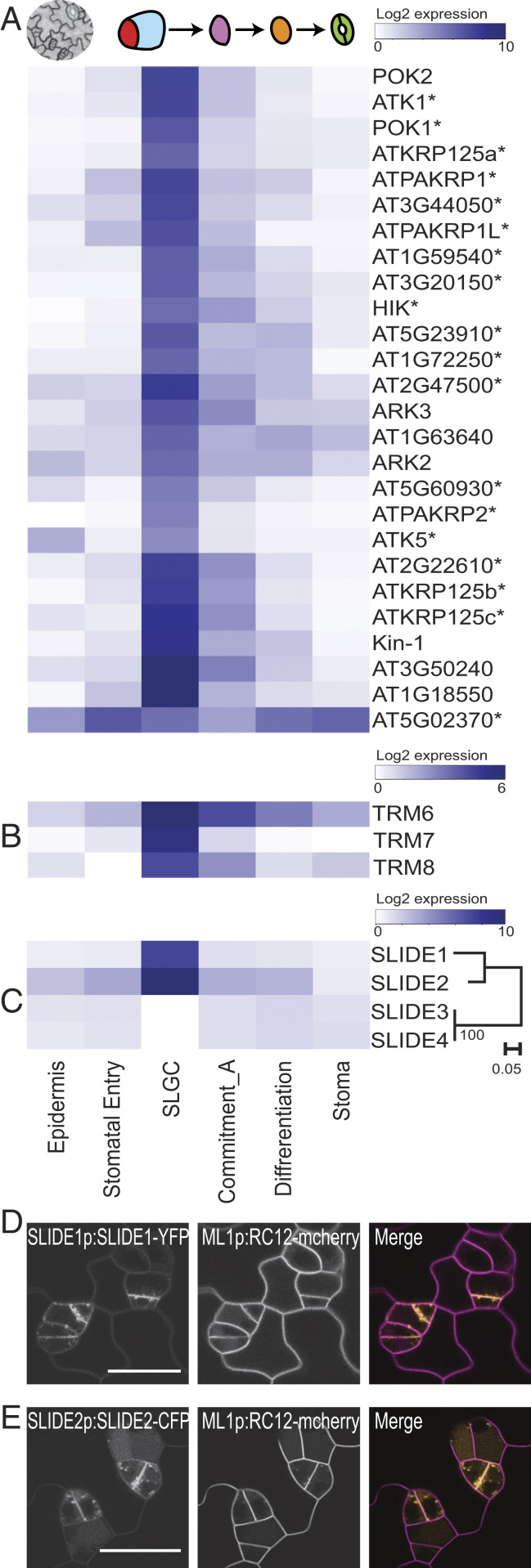
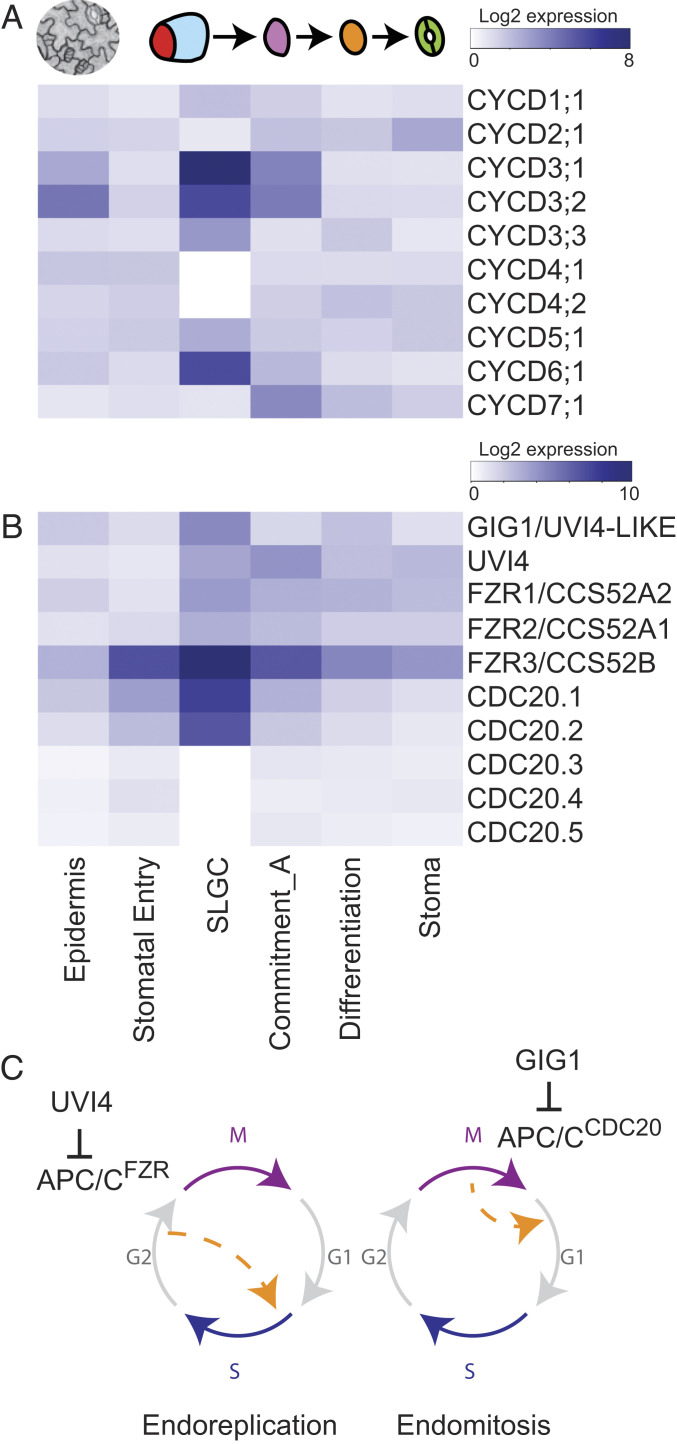
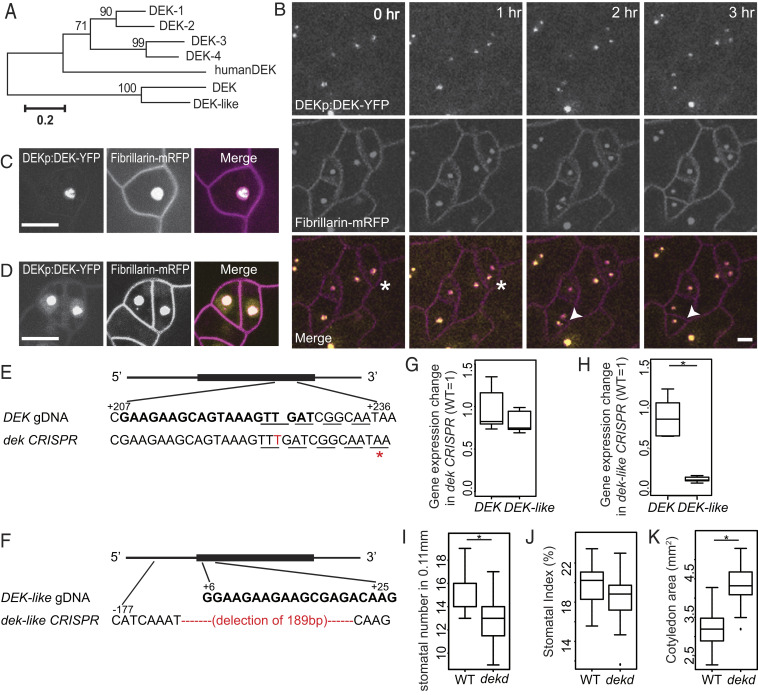
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
