

A Chlamydia trachomatis VD1-MOMP vaccine elicits cross-neutralizing and protective antibodies against C/C-related complex serovars
- PMID: 33875654
- PMCID: PMC8055873
- DOI: 10.1038/s41541-021-00312-9
A Chlamydia trachomatis VD1-MOMP vaccine elicits cross-neutralizing and protective antibodies against C/C-related complex serovars
Erratum in
-
Publisher Correction: A Chlamydia trachomatis VD1-MOMP vaccine elicits cross-neutralizing and protective antibodies against C/C-related complex serovars.NPJ Vaccines. 2022 Aug 30;7(1):104. doi: 10.1038/s41541-022-00521-w. NPJ Vaccines. 2022. PMID: 36042252 Free PMC article. No abstract available.
Abstract
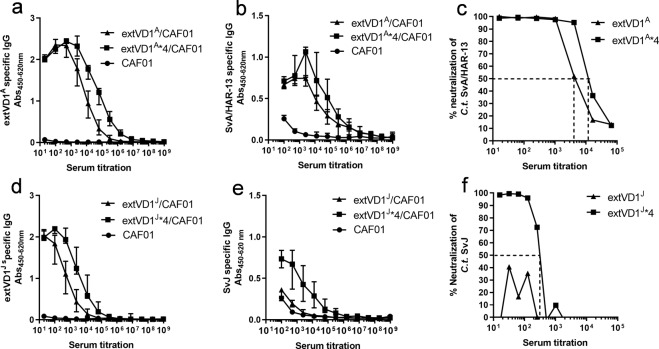
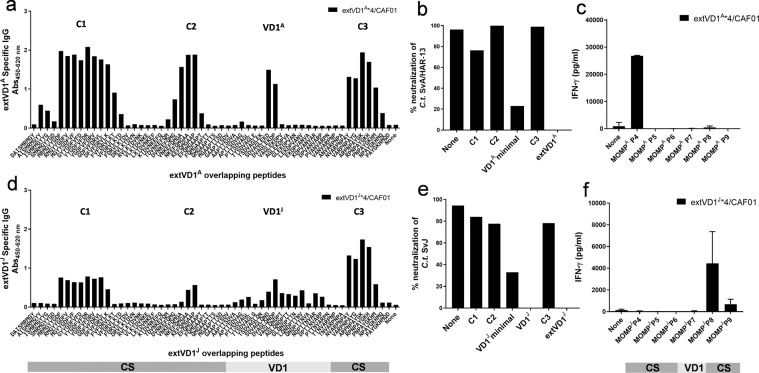
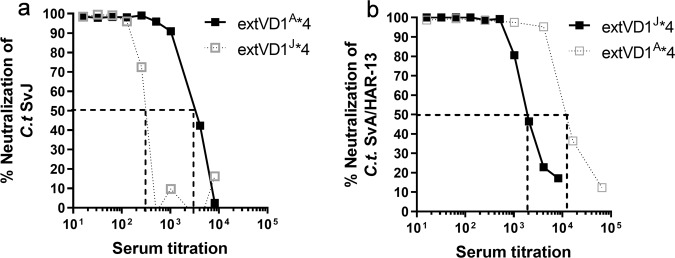
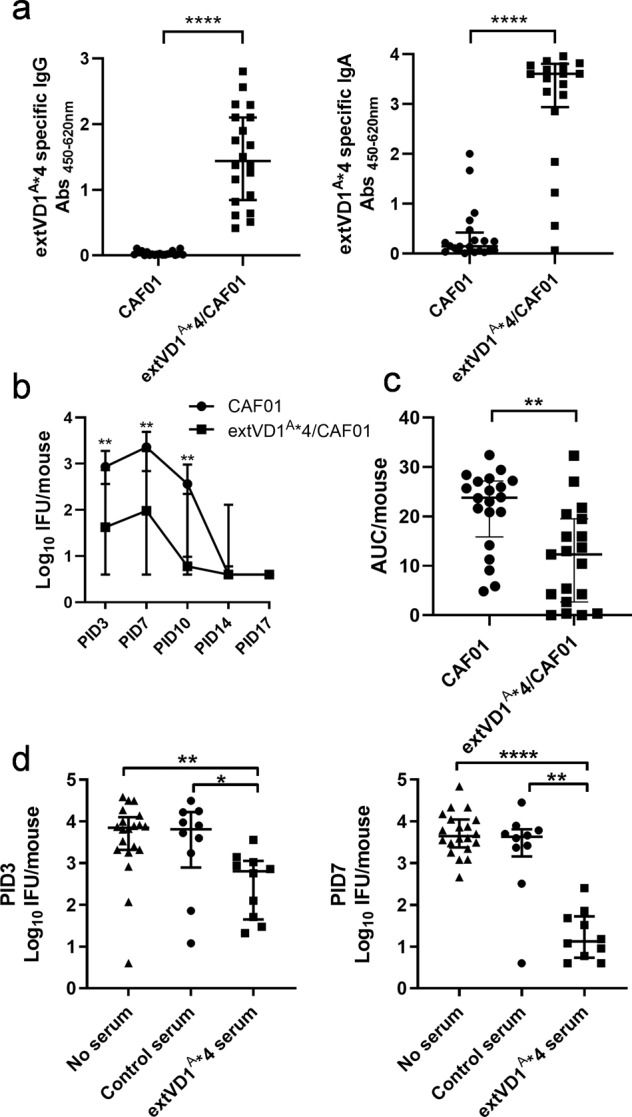
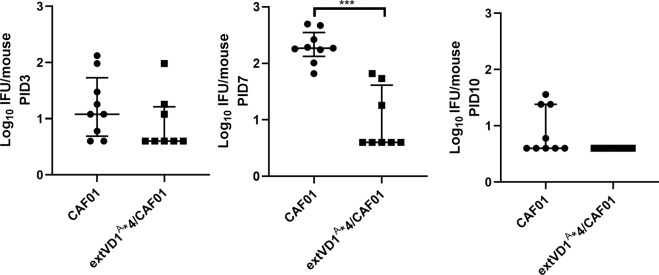
Ocular and urogenital infections with Chlamydia trachomatis (C.t.) are caused by a range of different serovars. The first C.t. vaccine in clinical development (CTH522/CAF®01) induced neutralizing antibodies directed to the variable domain 4 (VD4) region of major outer membrane protein (MOMP), covering predominantly B and intermediate groups of serovars. The VD1 region of MOMP contains neutralizing B-cell epitopes targeting serovars of the C and C-related complex. Using an immuno-repeat strategy, we extended the VD1 region of SvA and SvJ to include surrounding conserved segments, extVD1A and extVD1J, and repeated this region four times. The extVD1A*4 was most immunogenic with broad cross-surface and neutralizing reactivity against representative members of the C and C-related complex serovars. Importantly, in vitro results for extVD1A*4 translated into in vivo biological effects, demonstrated by in vivo neutralization of SvA and protection/cross-protection against intravaginal challenge with both SvA and the heterologous SvIa strain.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. A.W.O., I.R., P.A., and F.F. are co-inventors on a patent application relating to
Figures

Similar articles
-
Human antibody signatures towards the Chlamydia trachomatis major outer membrane protein after natural infection and vaccination.EBioMedicine. 2024 Jun;104:105140. doi: 10.1016/j.ebiom.2024.105140. Epub 2024 May 13. EBioMedicine. 2024. PMID: 38744110 Free PMC article.
-
Structural Assessment of Chlamydia trachomatis Major Outer Membrane Protein (MOMP)-Derived Vaccine Antigens and Immunological Profiling in Mice with Different Genetic Backgrounds.Vaccines (Basel). 2024 Jul 18;12(7):789. doi: 10.3390/vaccines12070789. Vaccines (Basel). 2024. PMID: 39066427 Free PMC article.
-
Protection Against Chlamydia trachomatis Infection and Upper Genital Tract Pathological Changes by Vaccine-Promoted Neutralizing Antibodies Directed to the VD4 of the Major Outer Membrane Protein.J Infect Dis. 2015 Sep 15;212(6):978-89. doi: 10.1093/infdis/jiv137. Epub 2015 Mar 6. J Infect Dis. 2015. PMID: 25748320
-
Publisher Correction: A Chlamydia trachomatis VD1-MOMP vaccine elicits cross-neutralizing and protective antibodies against C/C-related complex serovars.NPJ Vaccines. 2022 Aug 30;7(1):104. doi: 10.1038/s41541-022-00521-w. NPJ Vaccines. 2022. PMID: 36042252 Free PMC article. No abstract available.
-
Antibodies to Variable Domain 4 Linear Epitopes of the Chlamydia trachomatis Major Outer Membrane Protein Are Not Associated with Chlamydia Resolution or Reinfection in Women.mSphere. 2020 Sep 23;5(5):e00654-20. doi: 10.1128/mSphere.00654-20. mSphere. 2020. PMID: 32968007 Free PMC article.
Cited by
-
Viral Vector-Based Chlamydia trachomatis Vaccines Encoding CTH522 Induce Distinct Immune Responses in C57BL/6J and HLA Transgenic Mice.Vaccines (Basel). 2024 Aug 22;12(8):944. doi: 10.3390/vaccines12080944. Vaccines (Basel). 2024. PMID: 39204067 Free PMC article.
-
Heterologous prime-boost vaccination based on Polymorphic protein D protects against intravaginal Chlamydia trachomatis infection in mice.Sci Rep. 2022 Apr 22;12(1):6664. doi: 10.1038/s41598-022-10633-x. Sci Rep. 2022. PMID: 35459778 Free PMC article.
-
Overexpression of the Bam Complex Improves the Production of Chlamydia trachomatis MOMP in the E. coli Outer Membrane.Int J Mol Sci. 2022 Jul 2;23(13):7393. doi: 10.3390/ijms23137393. Int J Mol Sci. 2022. PMID: 35806397 Free PMC article.
-
Advances in Chlamydia trachomatis Vaccination: Unveiling the Potential of Major Outer Membrane Protein Derivative Constructs.Microorganisms. 2024 Jun 13;12(6):1196. doi: 10.3390/microorganisms12061196. Microorganisms. 2024. PMID: 38930578 Free PMC article. Review.
-
A systems serology approach to the investigation of infection-induced antibody responses and protection in trachoma.Front Immunol. 2023 May 23;14:1178741. doi: 10.3389/fimmu.2023.1178741. eCollection 2023. Front Immunol. 2023. PMID: 37287960 Free PMC article.
References
-
- Olsen AW, Follmann F, Erneholm K, Rosenkrands I, Andersen P. Protection against Chlamydia trachomatis Infection and upper genital tract pathological changes by vaccine-promoted neutralizing antibodies directed to the VD4 of the major outer membrane protein. J. Infect. Dis. 2015;212:978–989. doi: 10.1093/infdis/jiv137. - DOI - PubMed
Grants and funding
- DFF-4004-00424/Sundhed og Sygdom, Det Frie Forskningsråd (Medical Sciences, Danish Council for Independent Research)
- DFF-4004-00424/Sundhed og Sygdom, Det Frie Forskningsråd (Medical Sciences, Danish Council for Independent Research)
- DFF-4004-00424/Sundhed og Sygdom, Det Frie Forskningsråd (Medical Sciences, Danish Council for Independent Research)
LinkOut - more resources
Full Text Sources
Other Literature Sources
