

Composition and function of ciliary inner-dynein-arm subunits studied in Chlamydomonas reinhardtii
- PMID: 33876572
- PMCID: PMC8217317
- DOI: 10.1002/cm.21662
Composition and function of ciliary inner-dynein-arm subunits studied in Chlamydomonas reinhardtii
Abstract
Motile cilia (also interchangeably called "flagella") are conserved organelles extending from the surface of many animal cells and play essential functions in eukaryotes, including cell motility and environmental sensing. Large motor complexes, the ciliary dyneins, are present on ciliary outer-doublet microtubules and drive movement of cilia. Ciliary dyneins are classified into two general types: the outer dynein arms (ODAs) and the inner dynein arms (IDAs). While ODAs are important for generation of force and regulation of ciliary beat frequency, IDAs are essential for control of the size and shape of the bend, features collectively referred to as waveform. Also, recent studies have revealed unexpected links between IDA components and human diseases. In spite of their importance, studies on IDAs have been difficult since they are very complex and composed for several types of IDA motors, each unique in composition and location in the axoneme. Thanks in part to genetic, biochemical, and structural analysis of Chlamydomonas reinhardtii, we are beginning to understand the organization and function of the ciliary IDAs. In this review, we summarize the composition of Chlamydomonas IDAs particularly focusing on each subunit, and discuss the assembly, conservation, and functional role(s) of these IDA subunits. Furthermore, we raise several additional questions/challenges regarding IDAs, and discuss future perspectives of IDA studies.
Keywords: Chlamydomonas; IDA; cilia; flagella; inner-arm dynein; motility; subunit.
© 2021 Wiley Periodicals LLC.
Conflict of interest statement
Conflict of Interest
The authors declare there is no conflict of interest in this work.
Figures
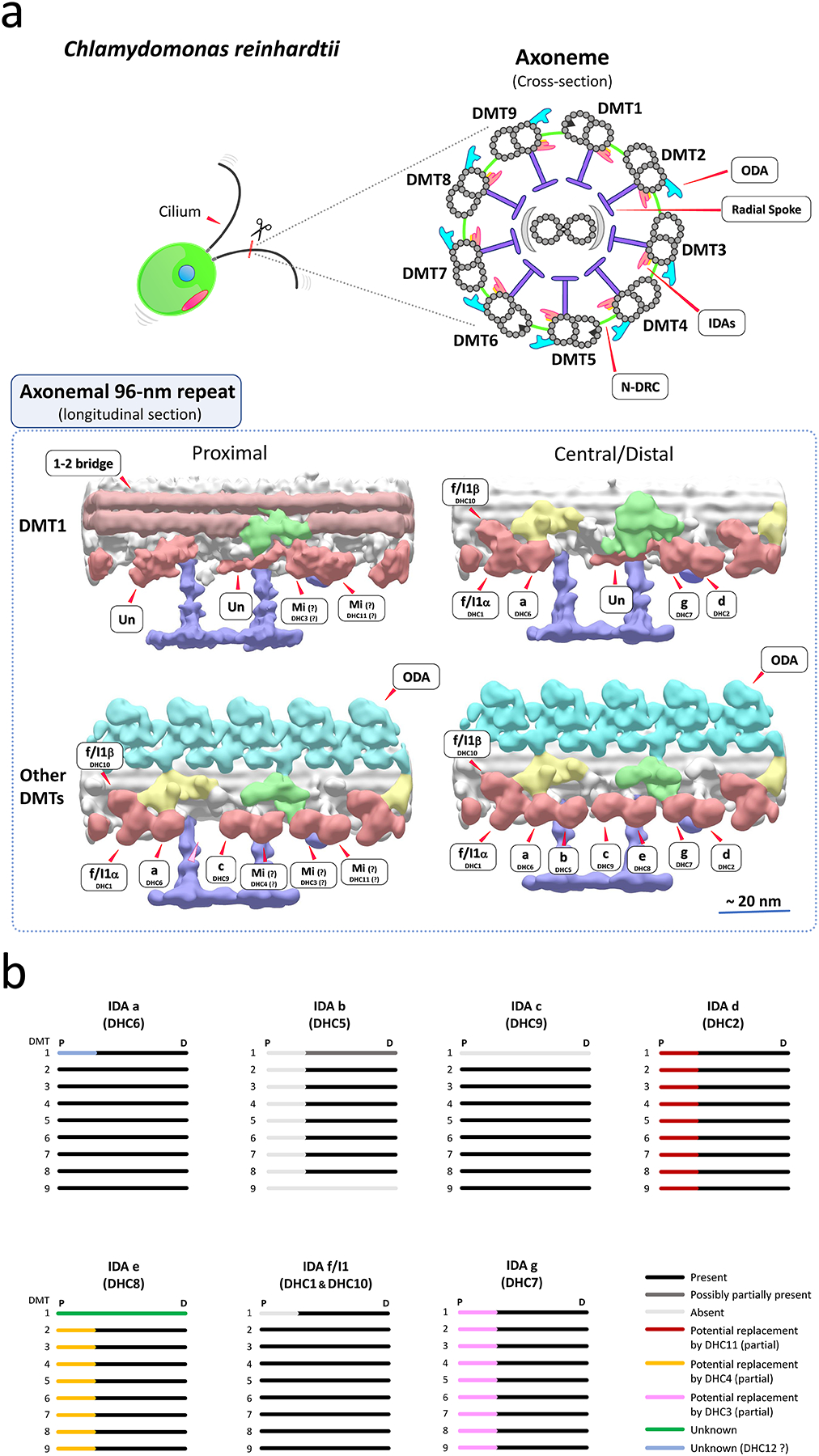
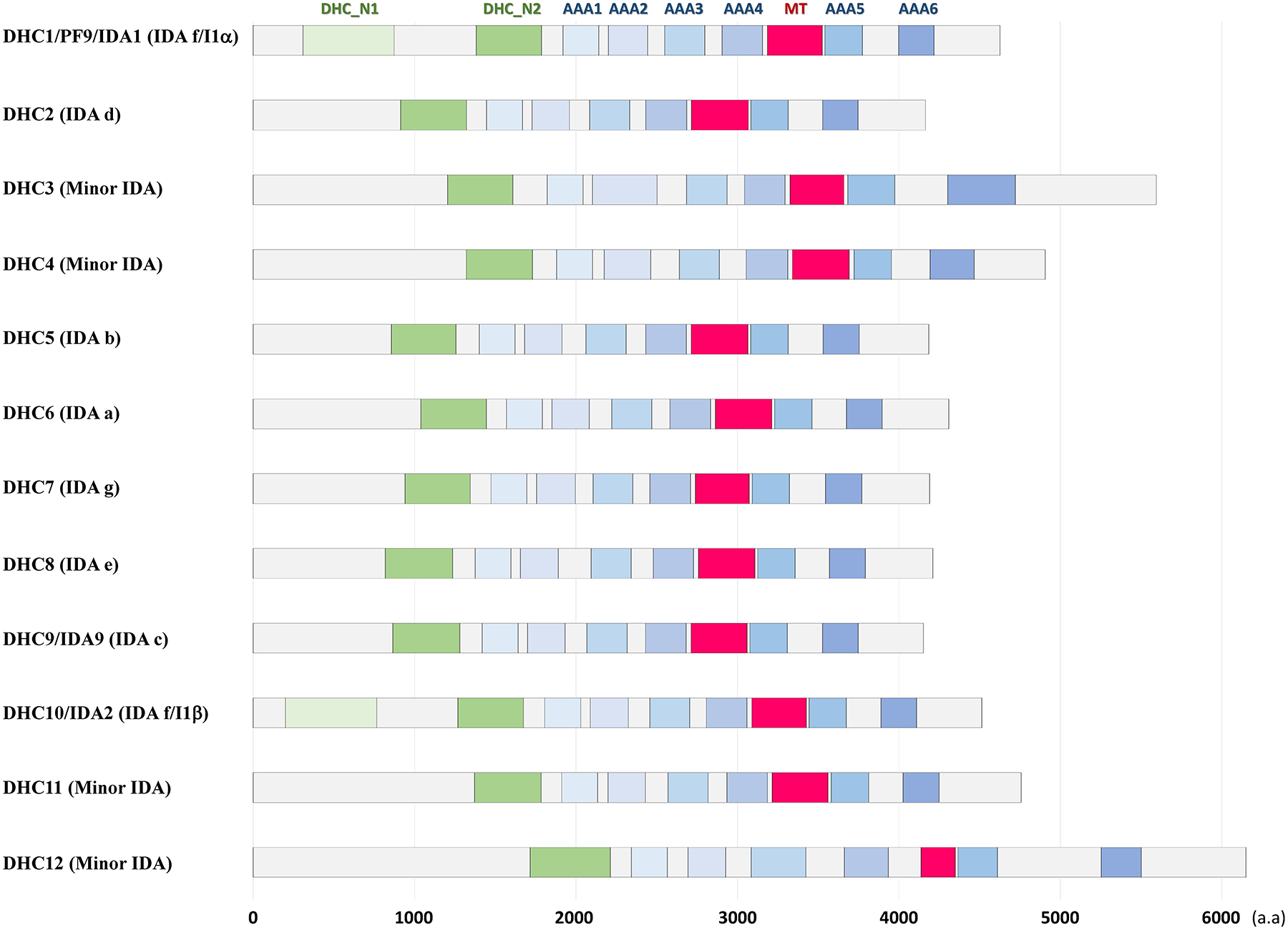
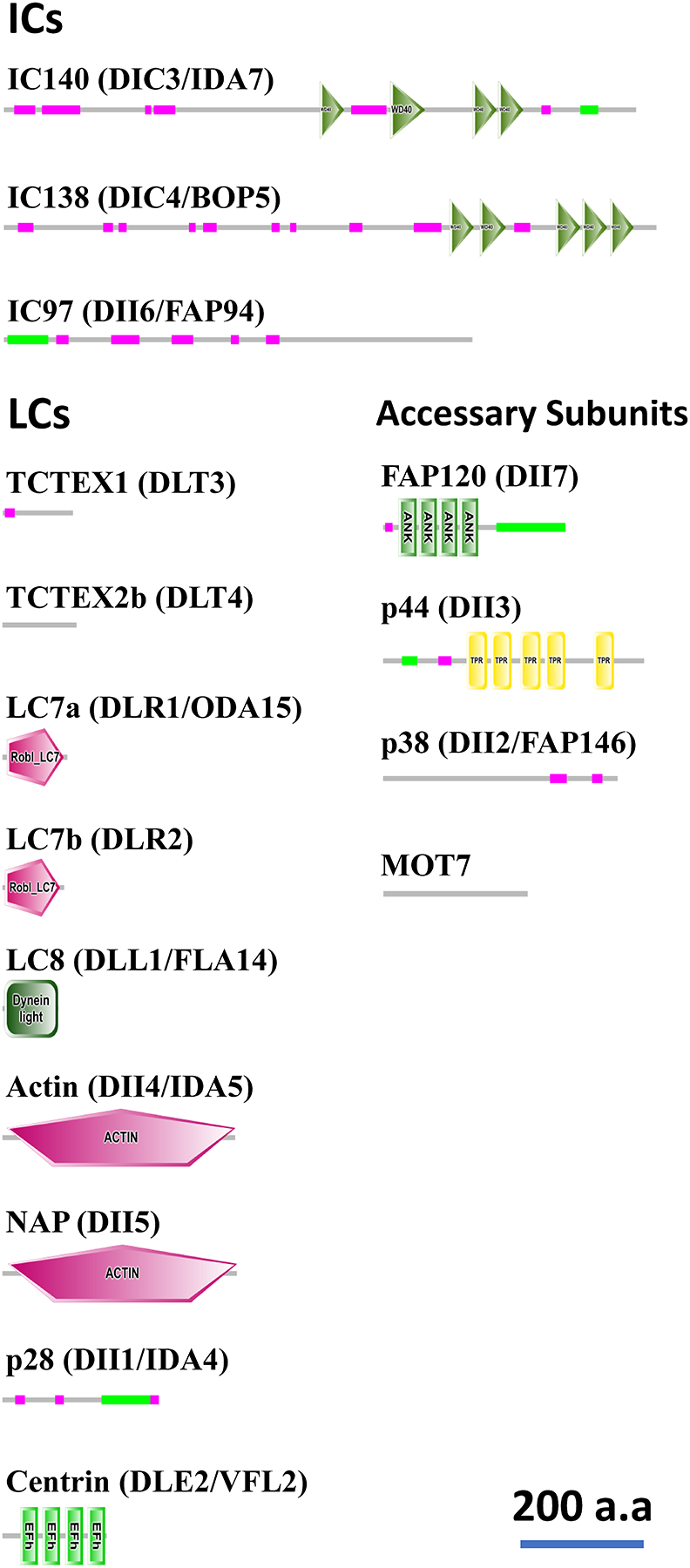
References
-
- Afzelius BA (1976). A human syndrome caused by immotile cilia. Science, 193(4250), 317–319. - PubMed
-
- Alford LM, Wirschell M, Yamamoto R, & Sale WS (2012). 11 - Control of Axonemal Inner Dynein Arms A2 - King, Stephen M. In Dyneins (pp. 312–335). Academic Press.
-
- Aravind Kumar M, Singh V, Naushad SM, Shanker U, & Lakshmi Narasu M (2018). Microarray-based SNP genotyping to identify genetic risk factors of triple-negative breast cancer (TNBC) in South Indian population. Mol Cell Biochem, 442(1–2), 1–10. - PubMed
-
- Barbar E (2008). Dynein light chain LC8 is a dimerization hub essential in diverse protein networks. Biochemistry, 47(2), 503–508. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
