

Inherited deficiency of stress granule ZNFX1 in patients with monocytosis and mycobacterial disease
- PMID: 33876776
- PMCID: PMC8053974
- DOI: 10.1073/pnas.2102804118
Inherited deficiency of stress granule ZNFX1 in patients with monocytosis and mycobacterial disease
Abstract
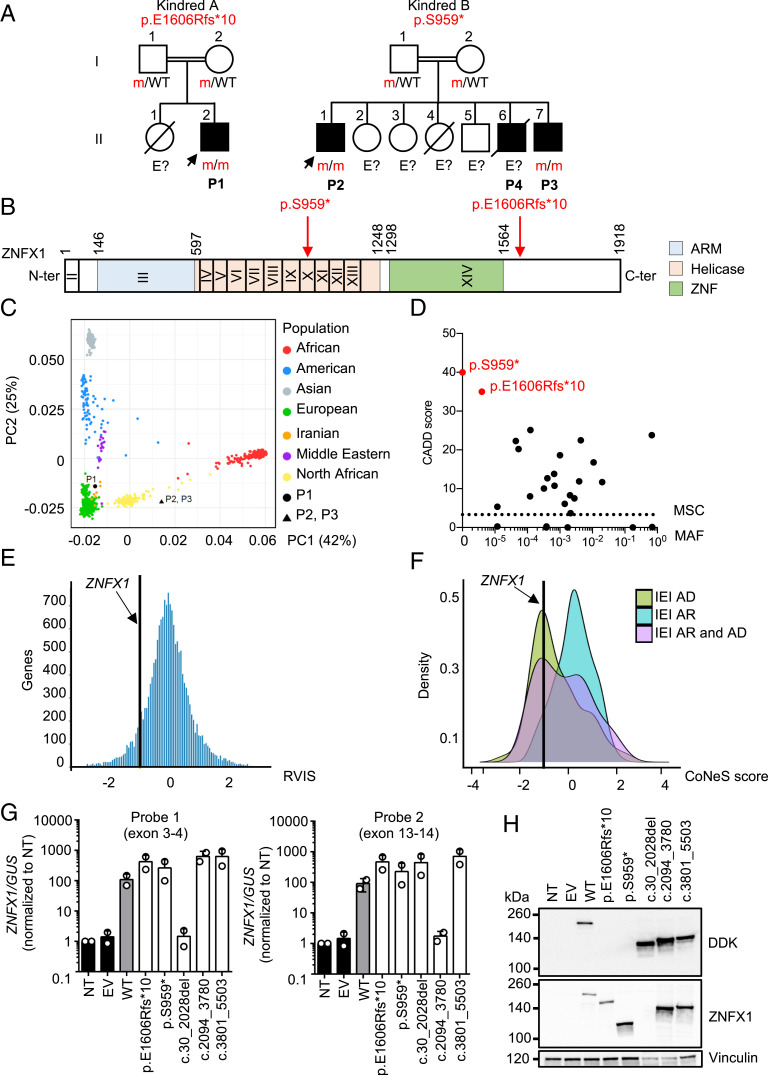
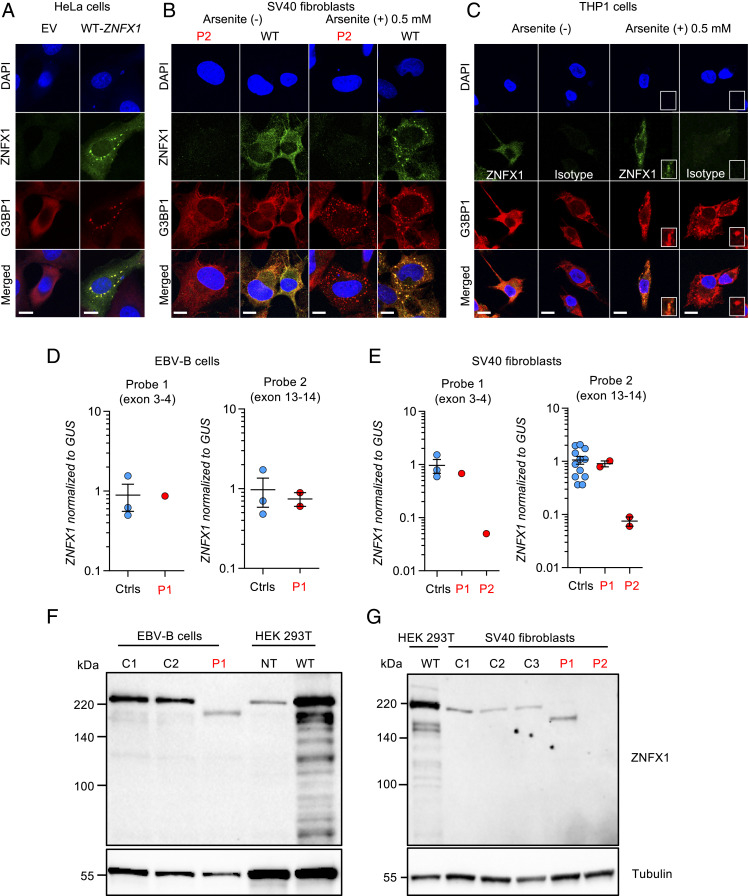
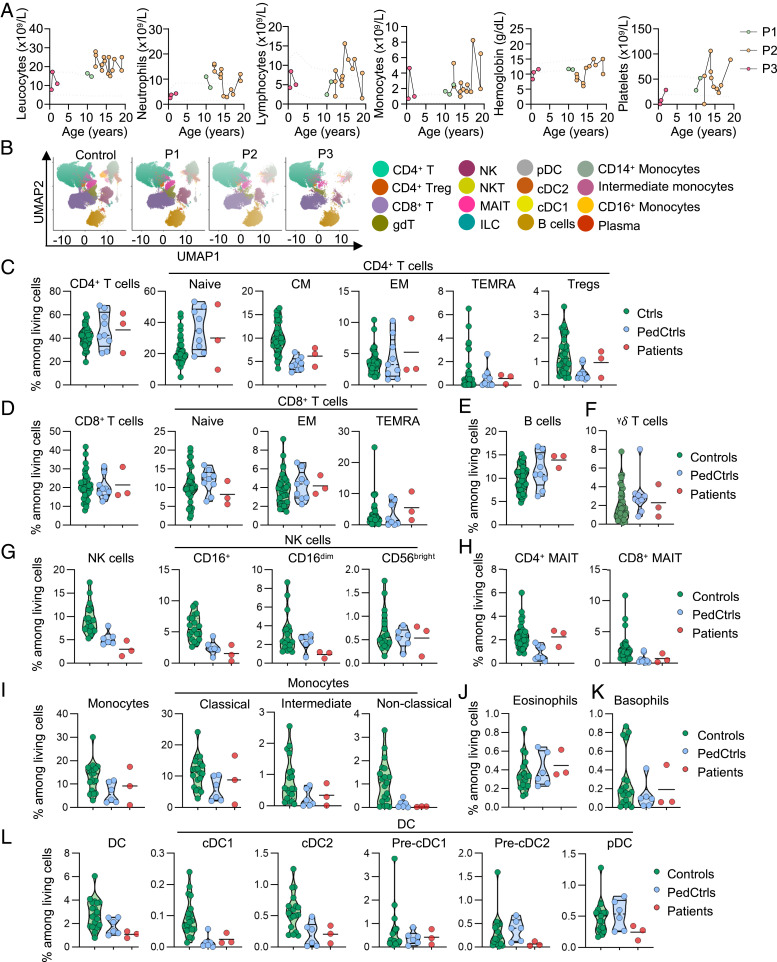
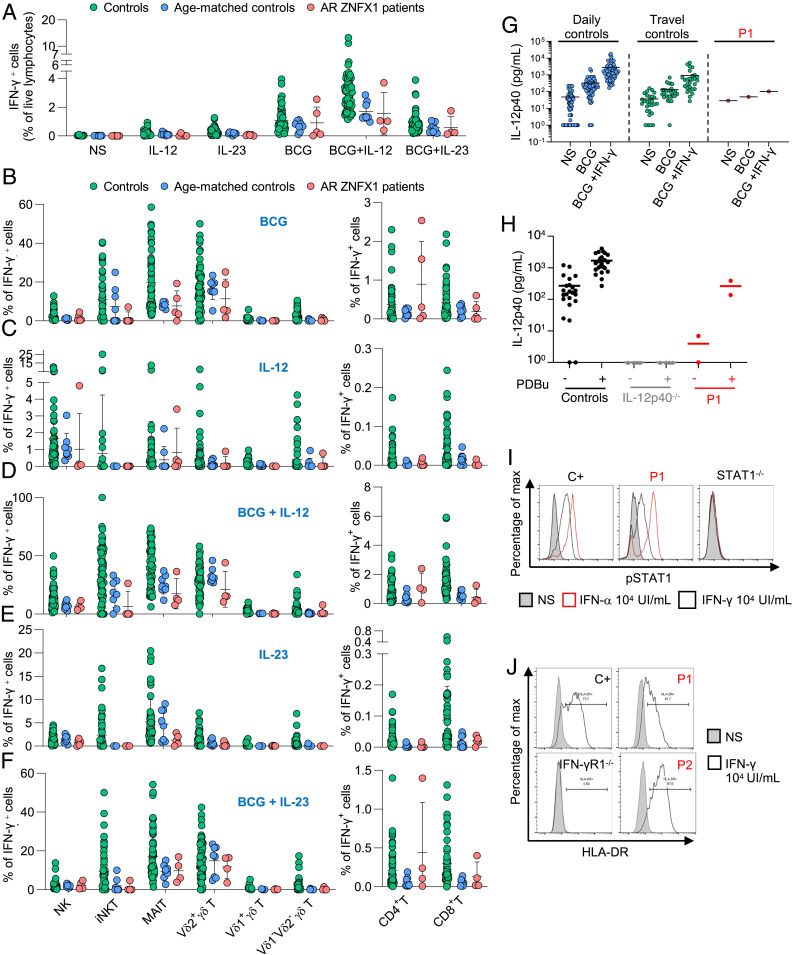
Human inborn errors of IFN-γ underlie mycobacterial disease, due to insufficient IFN-γ production by lymphoid cells, impaired myeloid cell responses to this cytokine, or both. We report four patients from two unrelated kindreds with intermittent monocytosis and mycobacterial disease, including bacillus Calmette-Guérin-osis and disseminated tuberculosis, and without any known inborn error of IFN-γ. The patients are homozygous for ZNFX1 variants (p.S959* and p.E1606Rfs*10) predicted to be loss of function (pLOF). There are no subjects homozygous for pLOF variants in public databases. ZNFX1 is a conserved and broadly expressed helicase, but its biology remains largely unknown. It is thought to act as a viral double-stranded RNA sensor in mice, but these patients do not suffer from severe viral illnesses. We analyze its subcellular localization upon overexpression in A549 and HeLa cell lines and upon stimulation of THP1 and fibroblastic cell lines. We find that this cytoplasmic protein can be recruited to or even induce stress granules. The endogenous ZNFX1 protein in cell lines of the patient homozygous for the p.E1606Rfs*10 variant is truncated, whereas ZNFX1 expression is abolished in cell lines from the patients with the p.S959* variant. Lymphocyte subsets are present at normal frequencies in these patients and produce IFN-γ normally. The hematopoietic and nonhematopoietic cells of the patients tested respond normally to IFN-γ. Our results indicate that human ZNFX1 is associated with stress granules and essential for both monocyte homeostasis and protective immunity to mycobacteria.
Keywords: ZNFX1; inborn error of immunity; inflammation; monocytosis; mycobacteria.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
