

Aging-dependent regulatory cells emerge in subcutaneous fat to inhibit adipogenesis
- PMID: 33878347
- PMCID: PMC8137669
- DOI: 10.1016/j.devcel.2021.03.026
Aging-dependent regulatory cells emerge in subcutaneous fat to inhibit adipogenesis
Abstract
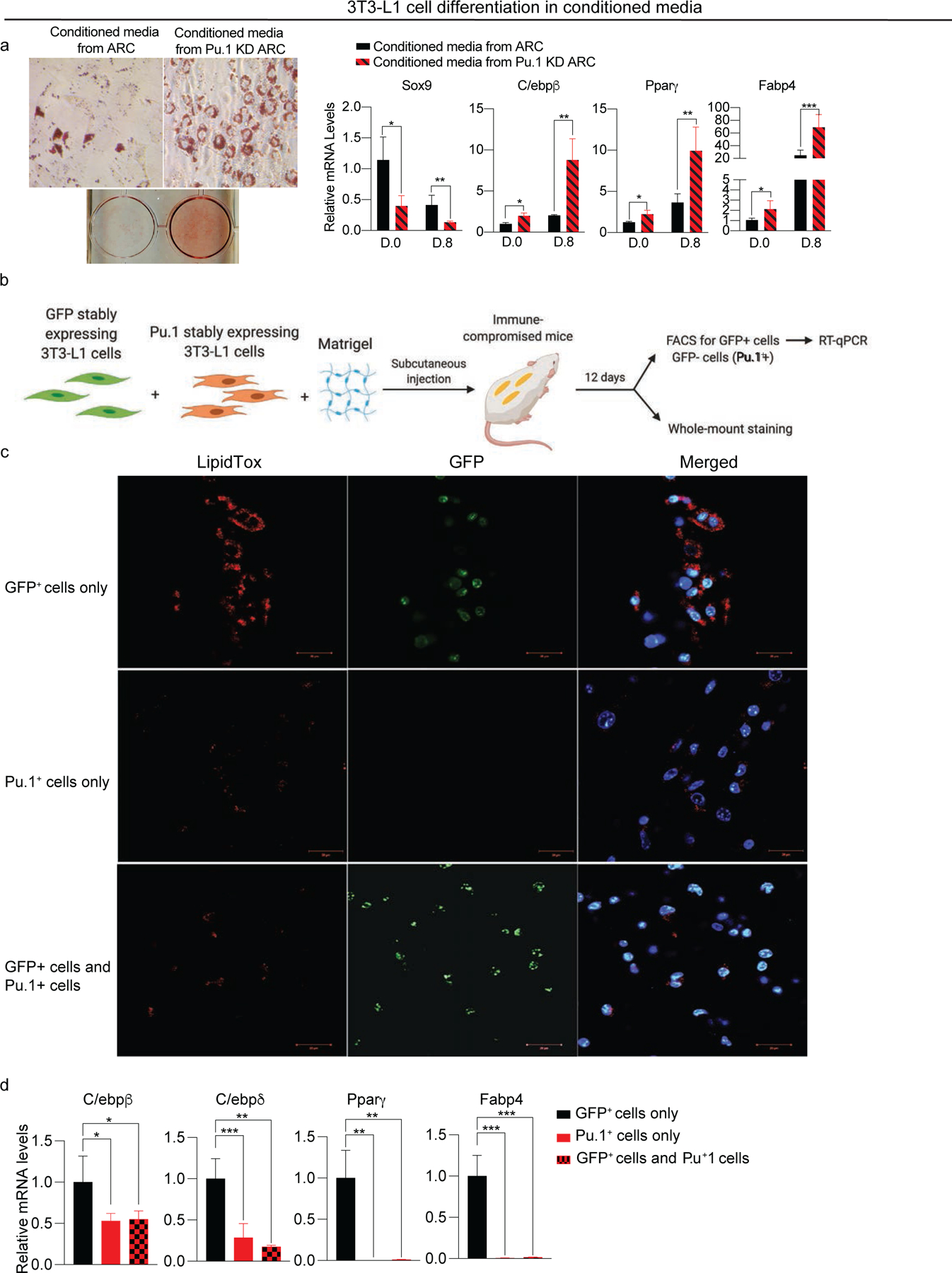
Adipose tissue mass and adiposity change throughout the lifespan. During aging, while visceral adipose tissue (VAT) tends to increase, peripheral subcutaneous adipose tissue (SAT) decreases significantly. Unlike VAT, which is linked to metabolic diseases, including type 2 diabetes, SAT has beneficial effects. However, the molecular details behind the aging-associated loss of SAT remain unclear. Here, by comparing scRNA-seq of total stromal vascular cells of SAT from young and aging mice, we identify an aging-dependent regulatory cell (ARC) population that emerges only in SAT of aged mice and humans. ARCs express adipose progenitor markers but lack adipogenic capacity; they secrete high levels of pro-inflammatory chemokines, including Ccl6, to inhibit proliferation and differentiation of neighboring adipose precursors. We also found Pu.1 to be a driving factor for ARC development. We identify an ARC population and its capacity to inhibit differentiation of neighboring adipose precursors, correlating with aging-associated loss of SAT.
Keywords: adipogenesis; adipose precursors; adipose tissue; aging; subcutaneous adipose tissue.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
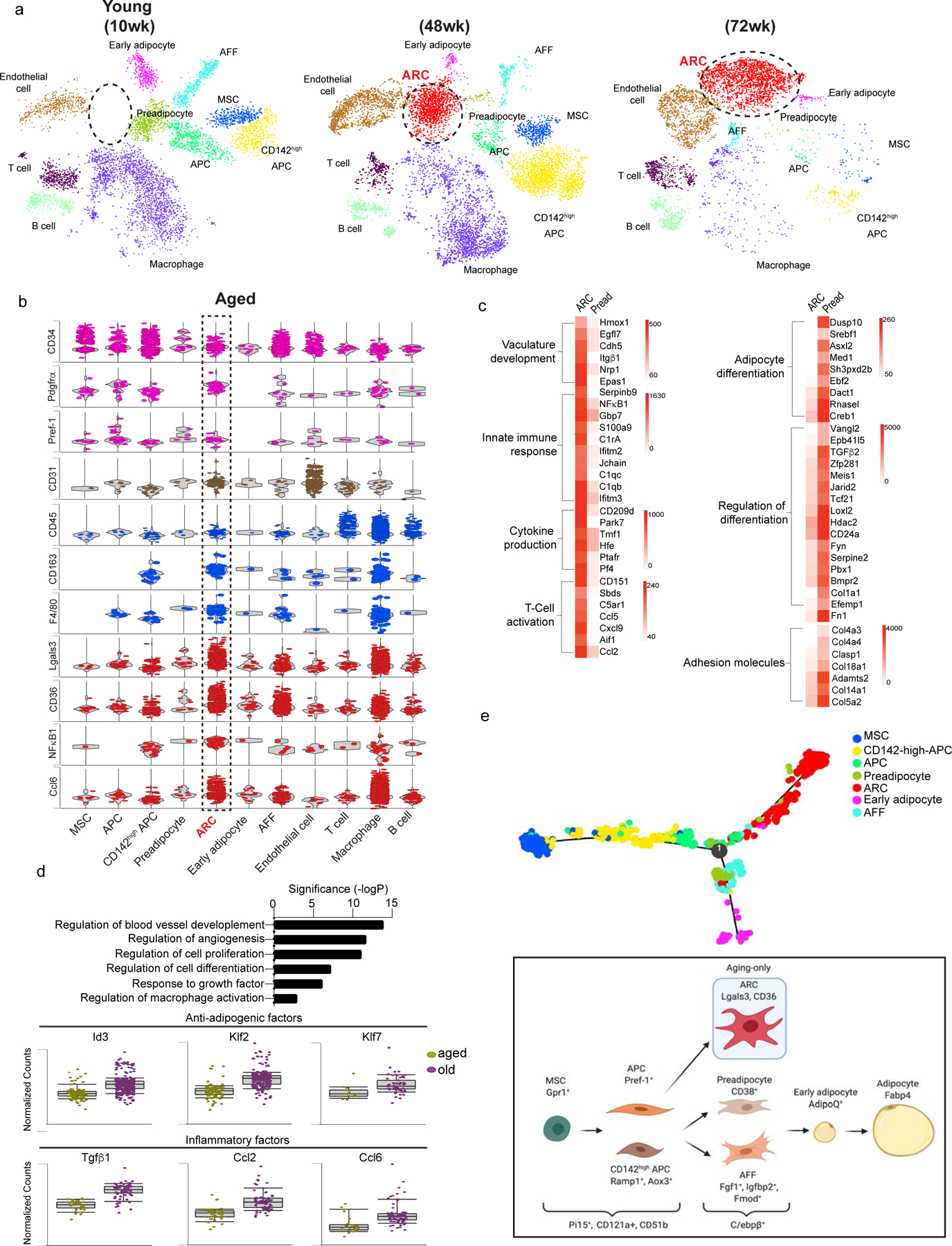
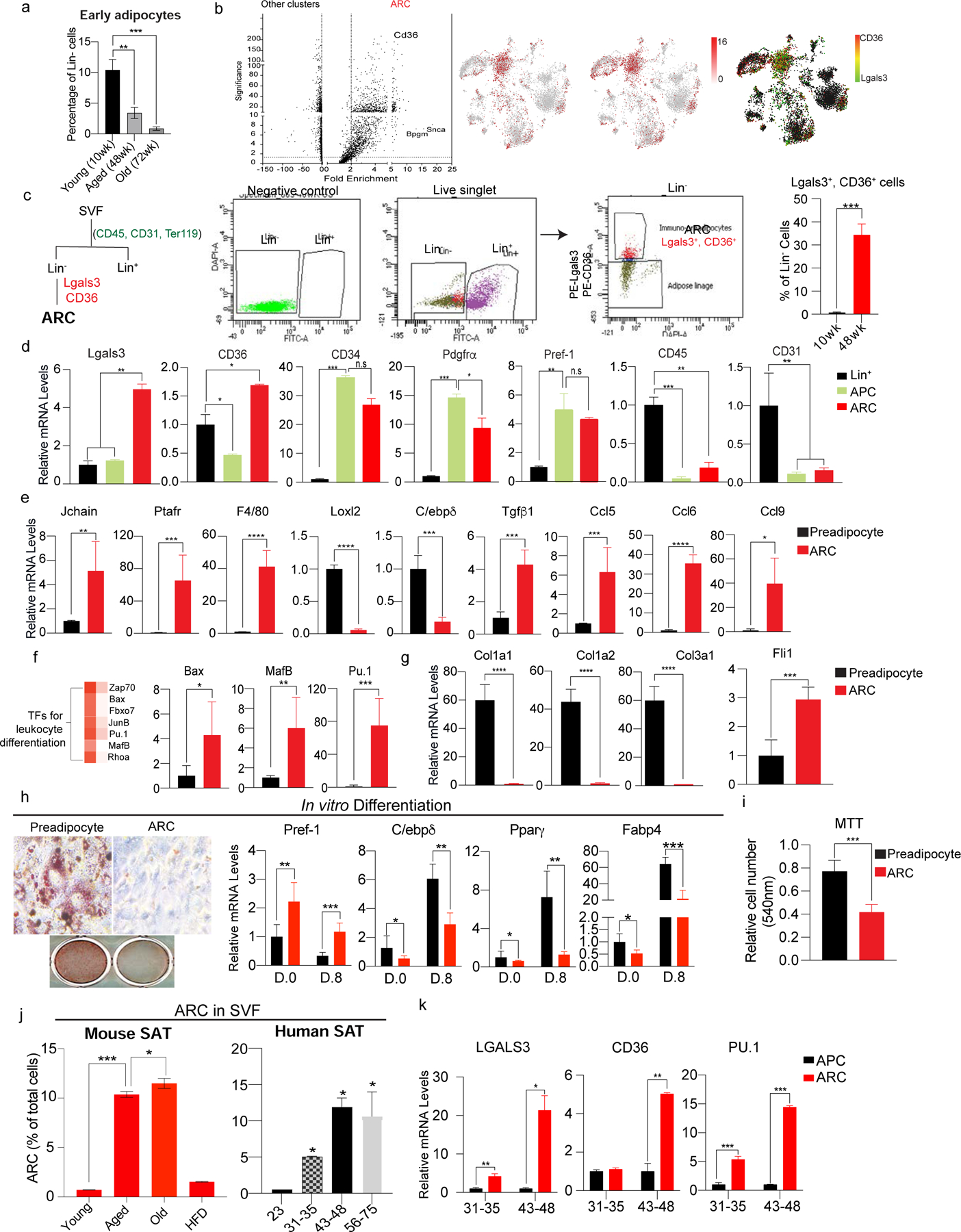
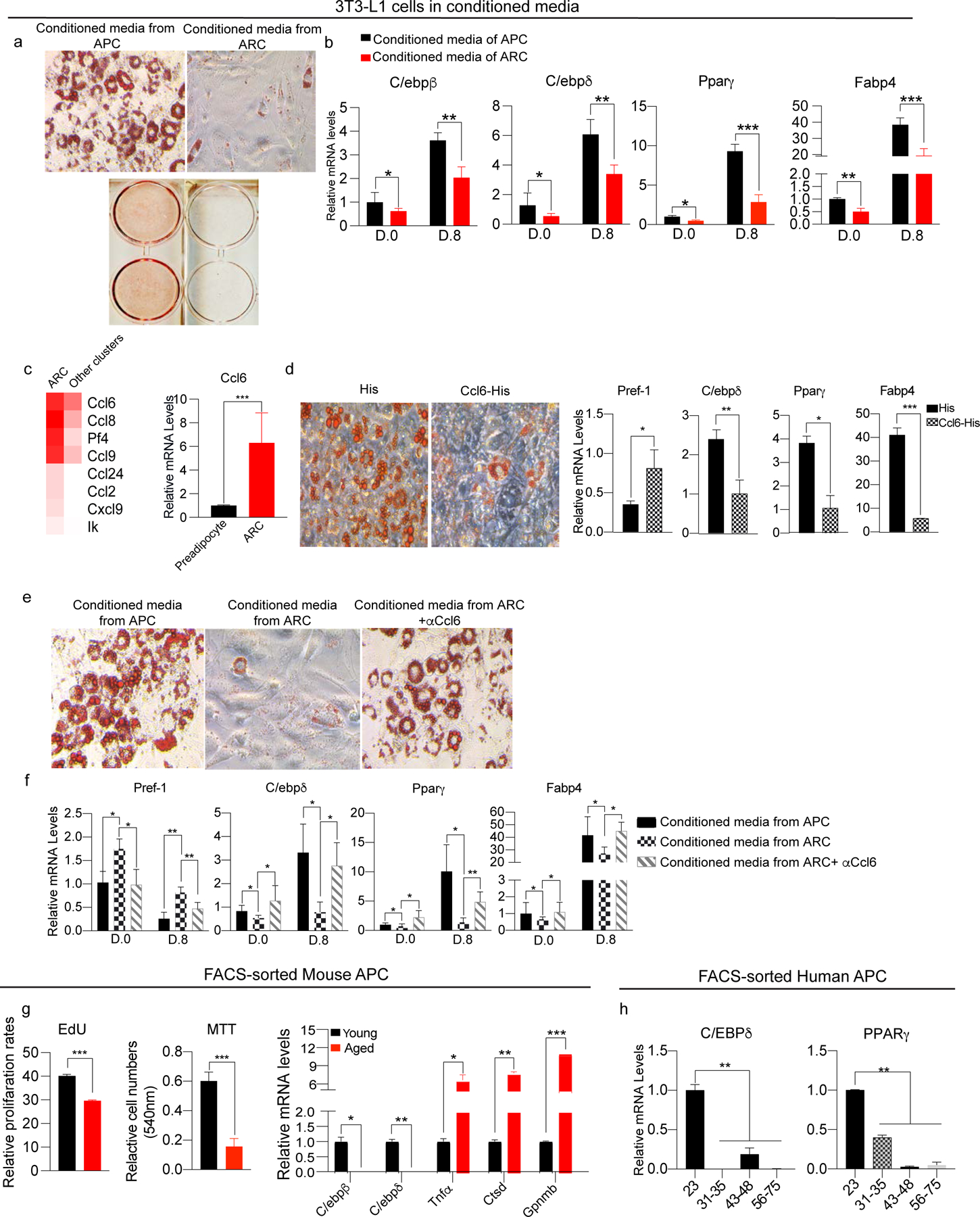
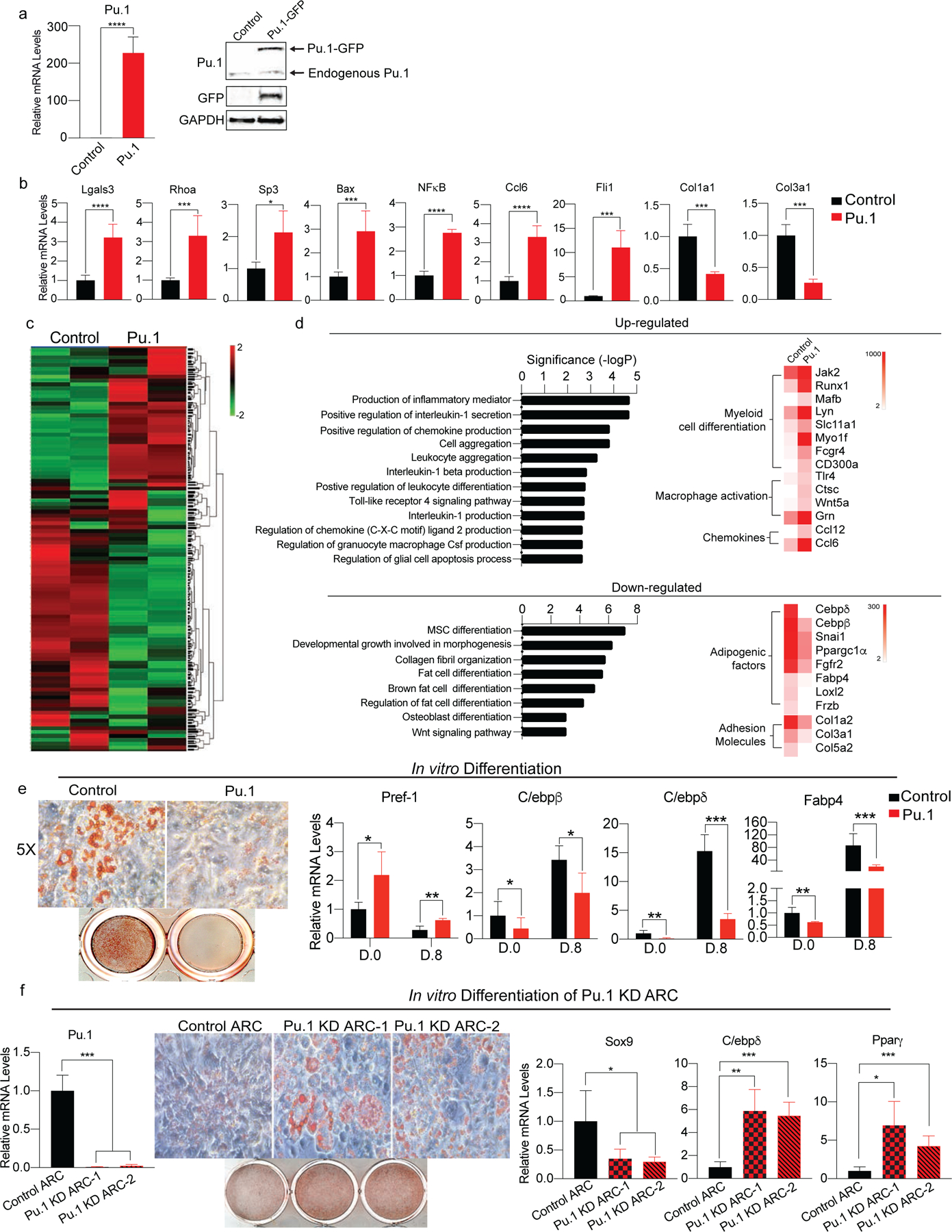
Comment in
-
Narrating the story ARC of adipose tissue aging.Dev Cell. 2021 May 17;56(10):1359-1360. doi: 10.1016/j.devcel.2021.04.020. Dev Cell. 2021. PMID: 34004148
References
-
- ALT EU, SENST C, MURTHY SN, SLAKEY DP, DUPIN CL, CHAFFIN AE, KADOWITZ PJ & IZADPANAH R 2012. Aging alters tissue resident mesenchymal stem cell properties. Stem Cell Res, 8, 215–25. - PubMed
-
- ARNER P 1995. Differences in lipolysis between human subcutaneous and omental adipose tissues. Ann Med, 27, 435–8. - PubMed
-
- BANERJEE SS, FEINBERG MW, WATANABE M, GRAY S, HASPEL RL, DENKINGER DJ, KAWAHARA R, HAUNER H & JAIN MK 2003. The Kruppel-like factor KLF2 inhibits peroxisome proliferator-activated receptor-gamma expression and adipogenesis. J Biol Chem, 278, 2581–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
