

Optimized polyepitope neoantigen DNA vaccines elicit neoantigen-specific immune responses in preclinical models and in clinical translation
- PMID: 33879241
- PMCID: PMC8059244
- DOI: 10.1186/s13073-021-00872-4
Optimized polyepitope neoantigen DNA vaccines elicit neoantigen-specific immune responses in preclinical models and in clinical translation
Abstract
Background: Preclinical studies and early clinical trials have shown that targeting cancer neoantigens is a promising approach towards the development of personalized cancer immunotherapies. DNA vaccines can be rapidly and efficiently manufactured and can integrate multiple neoantigens simultaneously. We therefore sought to optimize the design of polyepitope DNA vaccines and test optimized polyepitope neoantigen DNA vaccines in preclinical models and in clinical translation.
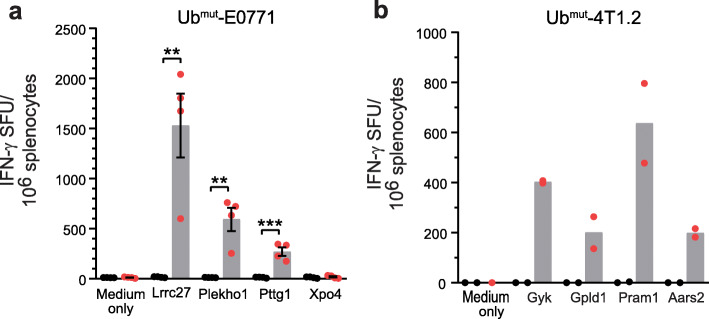
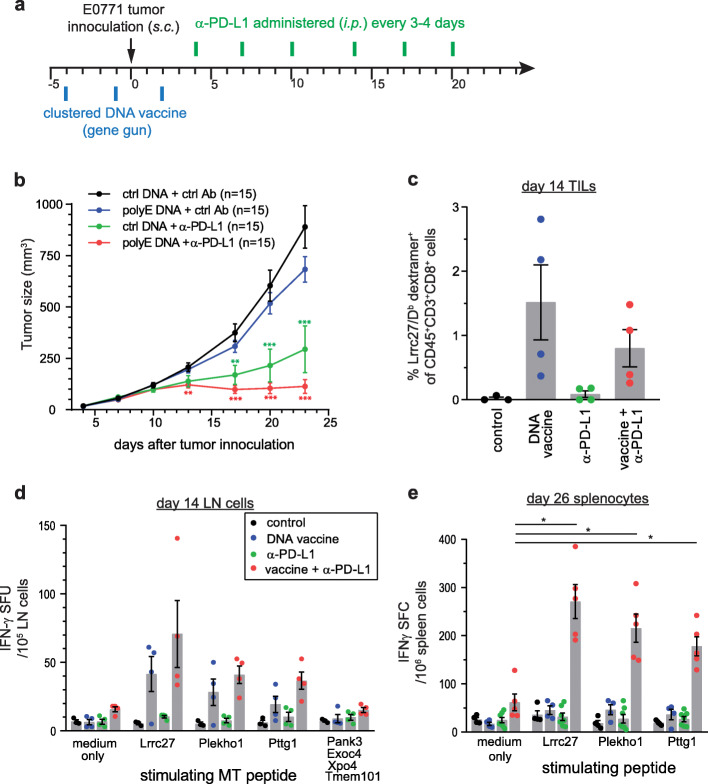
Methods: We developed and optimized a DNA vaccine platform to target multiple neoantigens. The polyepitope DNA vaccine platform was first optimized using model antigens in vitro and in vivo. We then identified neoantigens in preclinical breast cancer models through genome sequencing and in silico neoantigen prediction pipelines. Optimized polyepitope neoantigen DNA vaccines specific for the murine breast tumor E0771 and 4T1 were designed and their immunogenicity was tested in vivo. We also tested an optimized polyepitope neoantigen DNA vaccine in a patient with metastatic pancreatic neuroendocrine tumor.
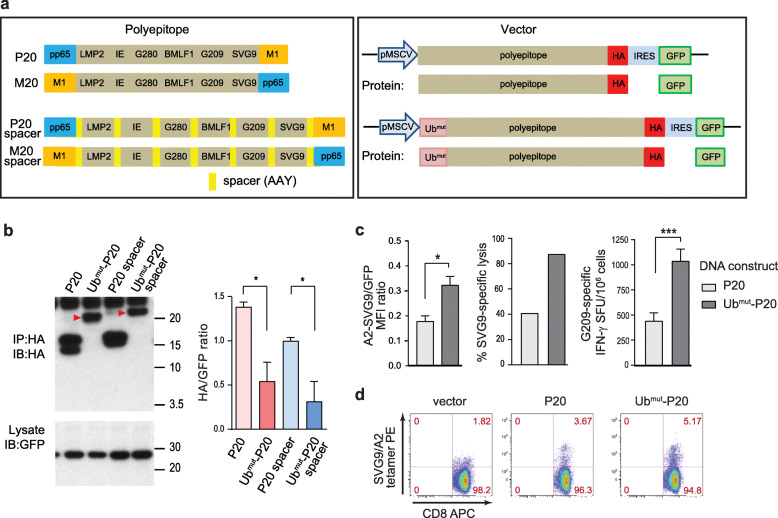
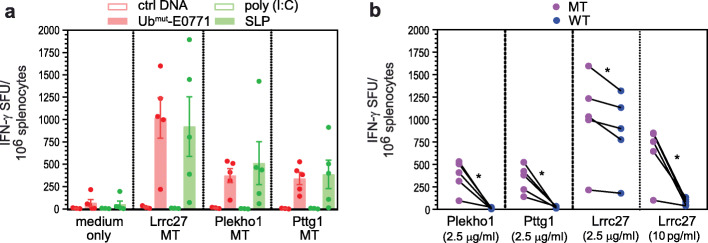
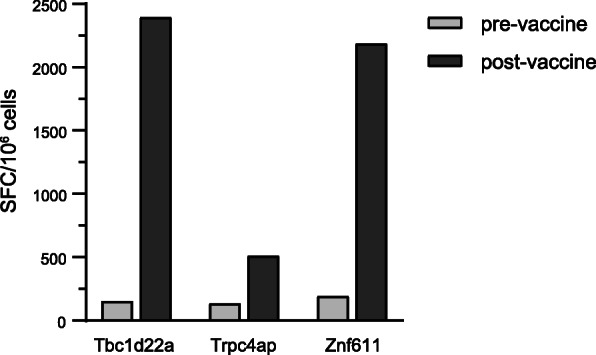
Results: Our data support an optimized polyepitope neoantigen DNA vaccine design encoding long (≥20-mer) epitopes with a mutant form of ubiquitin (Ubmut) fused to the N-terminus for antigen processing and presentation. Optimized polyepitope neoantigen DNA vaccines were immunogenic and generated robust neoantigen-specific immune responses in mice. The magnitude of immune responses generated by optimized polyepitope neoantigen DNA vaccines was similar to that of synthetic long peptide vaccines specific for the same neoantigens. When combined with immune checkpoint blockade therapy, optimized polyepitope neoantigen DNA vaccines were capable of inducing antitumor immunity in preclinical models. Immune monitoring data suggest that optimized polyepitope neoantigen DNA vaccines are capable of inducing neoantigen-specific T cell responses in a patient with metastatic pancreatic neuroendocrine tumor.
Conclusions: We have developed and optimized a novel polyepitope neoantigen DNA vaccine platform that can target multiple neoantigens and induce antitumor immune responses in preclinical models and neoantigen-specific responses in clinical translation.
Keywords: Breast cancer; Neoantigen; Polyepitope DNA vaccine.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Gubin MM, Zhang X, Schuster H, Caron E, Ward JP, Noguchi T, Ivanova Y, Hundal J, Arthur CD, Krebber WJ, Mulder GE, Toebes M, Vesely MD, Lam SSK, Korman AJ, Allison JP, Freeman GJ, Sharpe AH, Pearce EL, Schumacher TN, Aebersold R, Rammensee HG, Melief CJM, Mardis ER, Gillanders WE, Artyomov MN, Schreiber RD. Checkpoint blockade cancer immunotherapy targets tumour-specific mutant antigens. Nature. 2014;515(7528):577–581. doi: 10.1038/nature13988. - DOI - PMC - PubMed
-
- Matsushita H, Vesely MD, Koboldt DC, Rickert CG, Uppaluri R, Magrini VJ, Arthur CD, White JM, Chen YS, Shea LK, Hundal J, Wendl MC, Demeter R, Wylie T, Allison JP, Smyth MJ, Old LJ, Mardis ER, Schreiber RD. Cancer exome analysis reveals a T-cell-dependent mechanism of cancer immunoediting. Nature. 2012;482(7385):400–404. doi: 10.1038/nature10755. - DOI - PMC - PubMed
-
- Zhang X, Kim S, Hundal J, Herndon JM, Li S, Petti AA, Soysal SD, Li L, McLellan MD, Hoog J, Primeau T, Myers N, Vickery TL, Sturmoski M, Hagemann IS, Miller CA, Ellis MJ, Mardis ER, Hansen T, Fleming TP, Goedegebuure SP, Gillanders WE. Breast cancer neoantigens can induce CD8+ T-cell responses and antitumor immunity. Cancer Immunol Res. 2017;5(7):516–523. doi: 10.1158/2326-6066.CIR-16-0264. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
