

Host Immunity Alters Community Ecology and Stability of the Microbiome in a Caenorhabditis elegans Model
- PMID: 33879498
- PMCID: PMC8561663
- DOI: 10.1128/mSystems.00608-20
Host Immunity Alters Community Ecology and Stability of the Microbiome in a Caenorhabditis elegans Model
Abstract
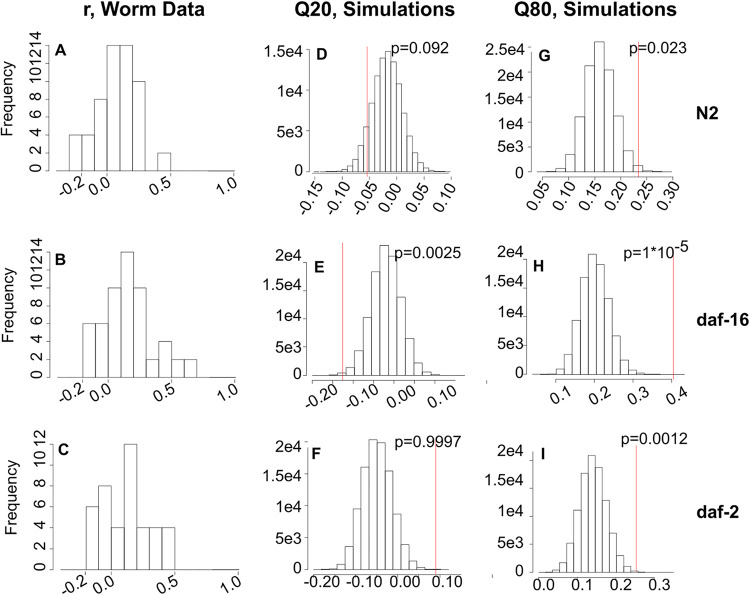
A growing body of data suggests that the microbiome of a species can vary considerably from individual to individual, but the reasons for this variation-and the consequences for the ecology of these communities-remain only partially explained. In mammals, the emerging picture is that the metabolic state and immune system status of the host affect the composition of the microbiome, but quantitative ecological microbiome studies are challenging to perform in higher organisms. Here, we show that these phenomena can be quantitatively analyzed in the tractable nematode host Caenorhabditis elegans Mutants in innate immunity, in particular the DAF-2/insulin growth factor (IGF) pathway, are shown to contain a microbiome that differs from that of wild-type nematodes. We analyzed the underlying basis of these differences from the perspective of community ecology by comparing experimental observations to the predictions of a neutral sampling model and concluded that fundamental differences in microbiome ecology underlie the observed differences in microbiome composition. We tested this hypothesis by introducing a minor perturbation into the colonization conditions, allowing us to assess stability of communities in different host strains. Our results show that altering host immunity changes the importance of interspecies interactions within the microbiome, resulting in differences in community composition and stability that emerge from these differences in host-microbe ecology.IMPORTANCE Here, we used a Caenorhabditis elegans microbiome model to demonstrate how genetic differences in innate immunity alter microbiome composition, diversity, and stability by changing the ecological processes that shape these communities. These results provide insight into the role of host genetics in controlling the ecology of the host-associated microbiota, resulting in differences in community composition, successional trajectories, and response to perturbation.
Keywords: Caenorhabditis elegans; immunocompromised hosts; microbial communities; microbial ecology.
Copyright © 2021 Taylor and Vega.
Figures




Similar articles
-
Gut-associated functions are favored during microbiome assembly across a major part of C. elegans life.mBio. 2024 May 8;15(5):e0001224. doi: 10.1128/mbio.00012-24. Epub 2024 Apr 18. mBio. 2024. PMID: 38634692 Free PMC article.
-
Natural genetic variation drives microbiome selection in the Caenorhabditis elegans gut.Curr Biol. 2021 Jun 21;31(12):2603-2618.e9. doi: 10.1016/j.cub.2021.04.046. Epub 2021 May 27. Curr Biol. 2021. PMID: 34048707 Free PMC article.
-
The native microbiome of the nematode Caenorhabditis elegans: gateway to a new host-microbiome model.BMC Biol. 2016 May 9;14:38. doi: 10.1186/s12915-016-0258-1. BMC Biol. 2016. PMID: 27160191 Free PMC article.
-
Understanding the factors regulating host-microbiome interactions using Caenorhabditis elegans.Philos Trans R Soc Lond B Biol Sci. 2024 May 6;379(1901):20230059. doi: 10.1098/rstb.2023.0059. Epub 2024 Mar 18. Philos Trans R Soc Lond B Biol Sci. 2024. PMID: 38497260 Free PMC article. Review.
-
Caenorhabditis elegans as a Convenient Animal Model for Microbiome Studies.Int J Mol Sci. 2024 Jun 18;25(12):6670. doi: 10.3390/ijms25126670. Int J Mol Sci. 2024. PMID: 38928375 Free PMC article. Review.
Cited by
-
Evolutionary implications of host genetic control for engineering beneficial microbiomes.Curr Opin Syst Biol. 2023 Jun;34:None. doi: 10.1016/j.coisb.2023.100455. Curr Opin Syst Biol. 2023. PMID: 37287906 Free PMC article. Review.
-
The intricate triangular interaction between protective microbe, pathogen and host determines fitness of the metaorganism.Proc Biol Sci. 2023 Dec 6;290(2012):20232193. doi: 10.1098/rspb.2023.2193. Epub 2023 Dec 6. Proc Biol Sci. 2023. PMID: 38052248 Free PMC article.
-
Master of Puppets: How Microbiota Drive the Nematoda Ecology and Evolution?Ecol Evol. 2025 Aug 19;15(8):e71549. doi: 10.1002/ece3.71549. eCollection 2025 Aug. Ecol Evol. 2025. PMID: 40837528 Free PMC article. Review.
-
Distinct members of the C. elegans CeMbio reference microbiota exert cryptic virulence and infection protection.bioRxiv [Preprint]. 2023 Nov 5:2023.11.02.565327. doi: 10.1101/2023.11.02.565327. bioRxiv. 2023. Update in: Mol Microbiol. 2024 Sep;122(3):387-402. doi: 10.1111/mmi.15258. PMID: 37961109 Free PMC article. Updated. Preprint.
-
The Interplay between the Host Microbiome and Pathogenic Viral Infections.mBio. 2021 Dec 21;12(6):e0249621. doi: 10.1128/mBio.02496-21. Epub 2021 Nov 2. mBio. 2021. PMID: 34724817 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
