

Specific modulation of the root immune system by a community of commensal bacteria
- PMID: 33879573
- PMCID: PMC8072228
- DOI: 10.1073/pnas.2100678118
Specific modulation of the root immune system by a community of commensal bacteria
Abstract
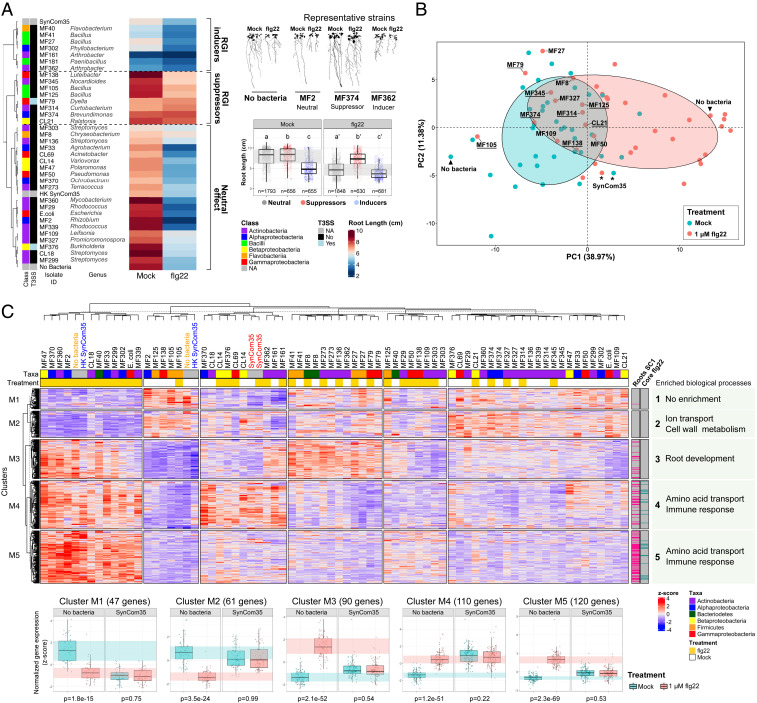
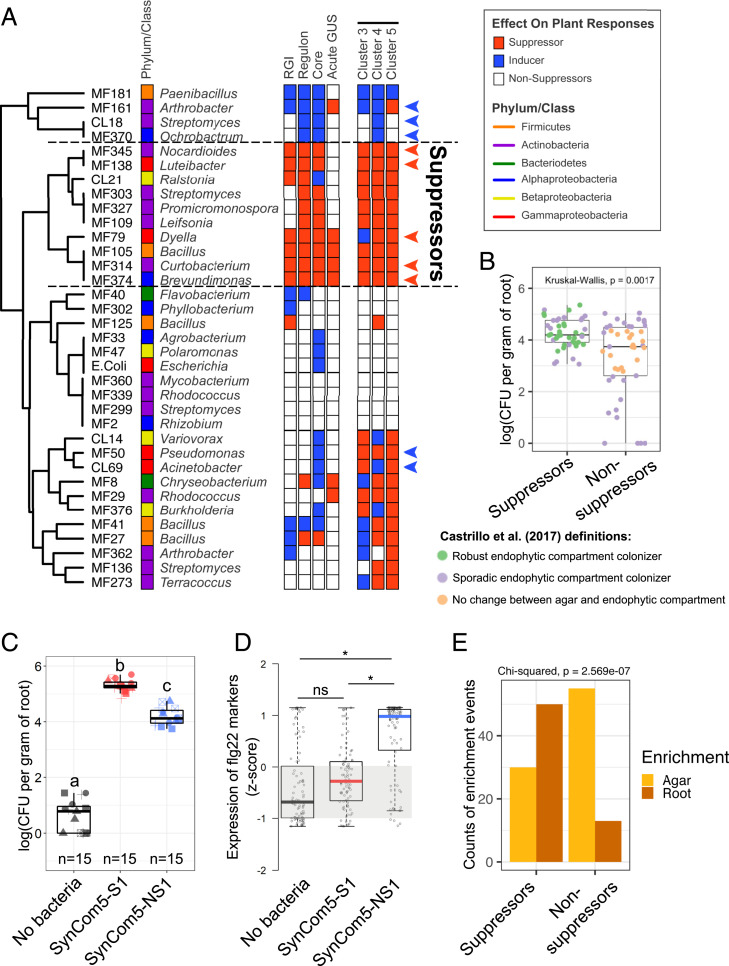
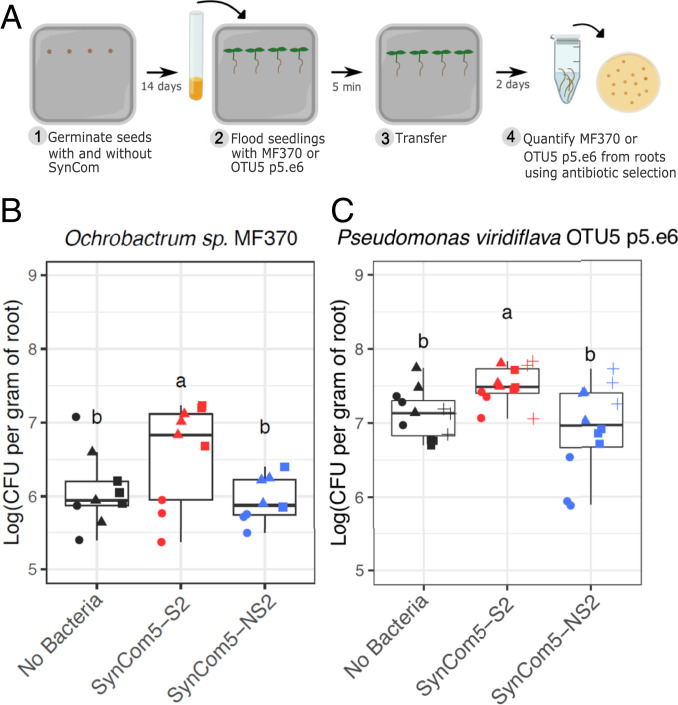
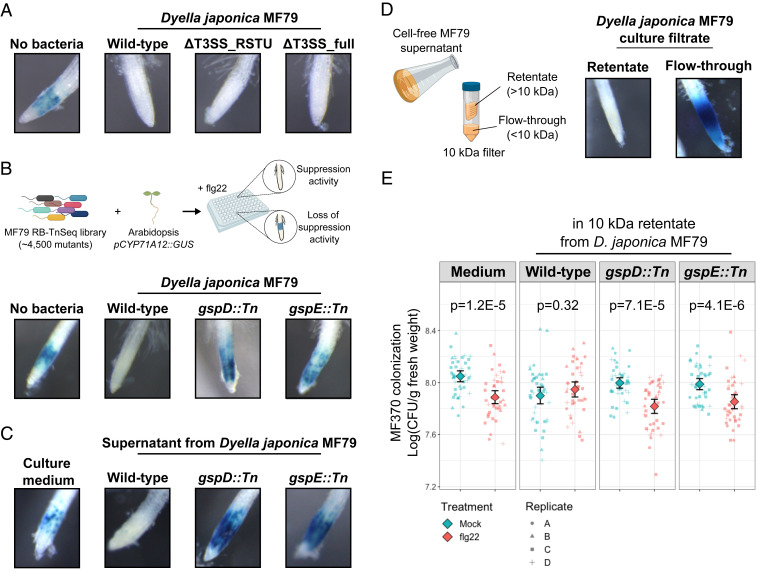
Plants have an innate immune system to fight off potential invaders that is based on the perception of nonself or modified-self molecules. Microbe-associated molecular patterns (MAMPs) are evolutionarily conserved microbial molecules whose extracellular detection by specific cell surface receptors initiates an array of biochemical responses collectively known as MAMP-triggered immunity (MTI). Well-characterized MAMPs include chitin, peptidoglycan, and flg22, a 22-amino acid epitope found in the major building block of the bacterial flagellum, FliC. The importance of MAMP detection by the plant immune system is underscored by the large diversity of strategies used by pathogens to interfere with MTI and that failure to do so is often associated with loss of virulence. Yet, whether or how MTI functions beyond pathogenic interactions is not well understood. Here we demonstrate that a community of root commensal bacteria modulates a specific and evolutionarily conserved sector of the Arabidopsis immune system. We identify a set of robust, taxonomically diverse MTI suppressor strains that are efficient root colonizers and, notably, can enhance the colonization capacity of other tested commensal bacteria. We highlight the importance of extracellular strategies for MTI suppression by showing that the type 2, not the type 3, secretion system is required for the immunomodulatory activity of one robust MTI suppressor. Our findings reveal that root colonization by commensals is controlled by MTI, which, in turn, can be selectively modulated by specific members of a representative bacterial root microbiota.
Keywords: MAMP; SynCom; flg22; plant immunity; root microbiome.
Conflict of interest statement
Competing interest statement: J.L.D. is a cofounder of, and shareholder in, AgBiome LLC, a corporation whose goal is to use plant-associated microbes to improve plant productivity.
Figures

References
-
- Fitzpatrick C. R., et al., The plant microbiome: From ecology to reductionism and beyond. Annu. Rev. Microbiol. 74, 81–100 (2020). - PubMed
-
- Teixeira P. J. P., Colaianni N. R., Fitzpatrick C. R., Dangl J. L., Beyond pathogens: Microbiota interactions with the plant immune system. Curr. Opin. Microbiol. 49, 7–17 (2019). - PubMed
-
- Hacquard S., Spaepen S., Garrido-Oter R., Schulze-Lefert P., Interplay between innate immunity and the plant microbiota. Annu. Rev. Phytopathol. 55, 565–589 (2017). - PubMed
-
- Lebeis S. L., et al., PLANT MICROBIOME. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 349, 860–864 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
