

In Vivo Antigen Expression Regulates CD4 T Cell Differentiation and Vaccine Efficacy against Mycobacterium tuberculosis Infection
- PMID: 33879592
- PMCID: PMC8092222
- DOI: 10.1128/mBio.00226-21
In Vivo Antigen Expression Regulates CD4 T Cell Differentiation and Vaccine Efficacy against Mycobacterium tuberculosis Infection
Abstract
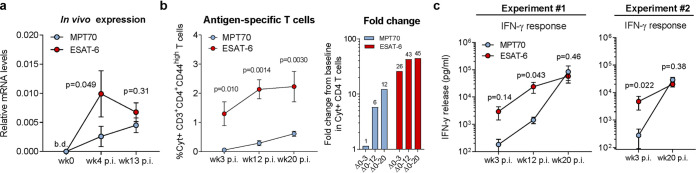
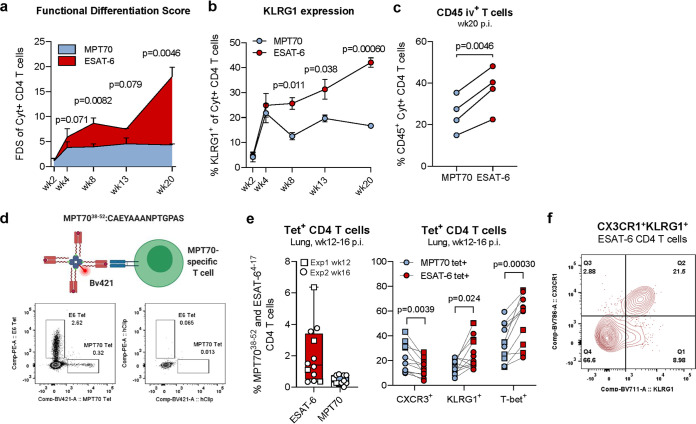
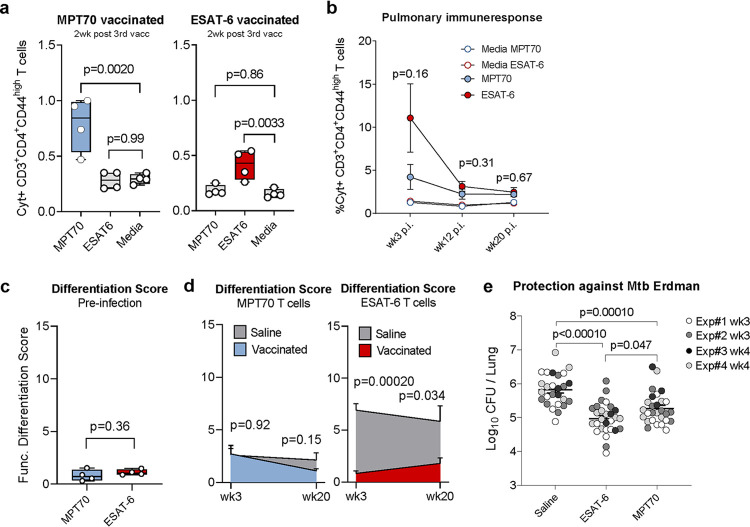
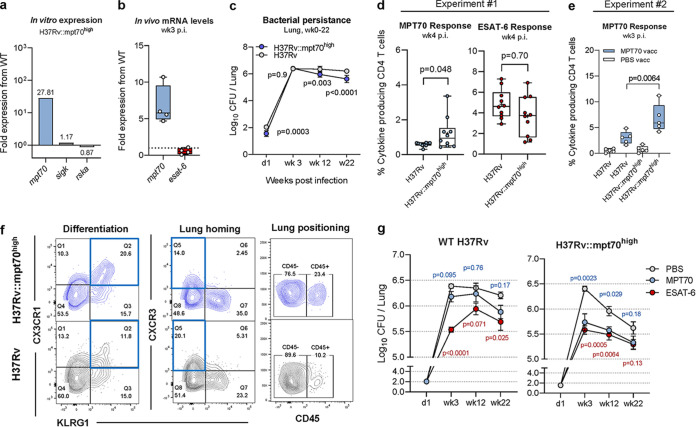
New vaccines are urgently needed against Mycobacterium tuberculosis (Mtb), which kills more than 1.4 million people each year. CD4 T cell differentiation is a key determinant of protective immunity against Mtb, but it is not fully understood how host-pathogen interactions shape individual antigen-specific T cell populations and their protective capacity. Here, we investigated the immunodominant Mtb antigen, MPT70, which is upregulated in response to gamma interferon (IFN-γ) or nutrient/oxygen deprivation of in vitro-infected macrophages. Using a murine aerosol infection model, we compared the in vivo expression kinetics of MPT70 to a constitutively expressed antigen, ESAT-6, and analyzed their corresponding CD4 T cell phenotype and vaccine protection. For wild-type Mtb, we found that in vivo expression of MPT70 was delayed compared to ESAT-6. This delayed expression was associated with induction of less differentiated MPT70-specific CD4 T cells but, compared to ESAT-6, also reduced protection after vaccination. In contrast, infection with an MPT70-overexpressing Mtb strain promoted highly differentiated KLRG1+CX3CR1+ CD4 T cells with limited lung-homing capacity. Importantly, this differentiated phenotype could be prevented by vaccination, and against the overexpressing strain, vaccination with MPT70 conferred protection similar to vaccination with ESAT-6. Together, our data indicate that high in vivo antigen expression drives T cells toward terminal differentiation and that targeted vaccination with adjuvanted protein can counteract this phenomenon by maintaining T cells in a protective less differentiated state. These observations shed new light on host-pathogen interactions and provide guidance on how future Mtb vaccines can be designed to tip the immune balance in favor of the host.IMPORTANCE Tuberculosis, caused by Mtb, constitutes a global health crisis of massive proportions and the impact of the current coronavirus disease 2019 (COVID-19) pandemic is expected to cause a rise in tuberculosis-related deaths. Improved vaccines are therefore needed more than ever, but a lack of knowledge on protective immunity hampers their development. The present study shows that constitutively expressed antigens with high availability drive highly differentiated CD4 T cells with diminished protective capacity, which could be a survival strategy by Mtb to evade T cell immunity against key antigens. We demonstrate that immunization with such antigens can counteract this phenomenon by maintaining antigen-specific T cells in a state of low differentiation. Future vaccine strategies should therefore explore combinations of multiple highly expressed antigens and we suggest that T cell differentiation could be used as a readily measurable parameter to identify these in both preclinical and clinical studies.
Keywords: ESAT-6; MPT70; Mycobacterium tuberculosis; T cell differentiation; immunization; in vivo expression; vaccination.
Copyright © 2021 Clemmensen et al.
Figures
Update of
-
In vivo antigen expression regulates CD4 T cell differentiation and vaccine efficacy against Mycobacterium tuberculosis infection.bioRxiv [Preprint]. 2021 Feb 3:2021.02.02.429488. doi: 10.1101/2021.02.02.429488. bioRxiv. 2021. Update in: mBio. 2021 Apr 20;12(2):e00226-21. doi: 10.1128/mBio.00226-21. PMID: 33564764 Free PMC article. Updated. Preprint.
References
-
- World Health Organization. 2020. Global tuberculosis report 2020. World Health Organization, Geneva, Switzerland.
-
- Sharma D, Bose A, Shakila H, Das TK, Tyagi JS, Ramanathan VD. 2006. Expression of mycobacterial cell division protein, FtsZ, and dormancy proteins, DevR and Acr, within lung granulomas throughout guinea pig infection. FEMS Immunol Med Microbiol 48:329–336. doi: 10.1111/j.1574-695X.2006.00160.x. - DOI - PubMed
-
- Schnappinger D, Ehrt S, Voskuil MI, Liu Y, Mangan JA, Monahan IM, Dolganov G, Efron B, Butcher PD, Nathan C, Schoolnik GK. 2003. Transcriptional adaptation of Mycobacterium tuberculosis within macrophages: insights into the phagosomal environment. J Exp Med 198:693–704. doi: 10.1084/jem.20030846. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
