

Dimer Asymmetry and Light Activation Mechanism in Brucella Blue-Light Sensor Histidine Kinase
- PMID: 33879593
- PMCID: PMC8092228
- DOI: 10.1128/mBio.00264-21
Dimer Asymmetry and Light Activation Mechanism in Brucella Blue-Light Sensor Histidine Kinase
Abstract
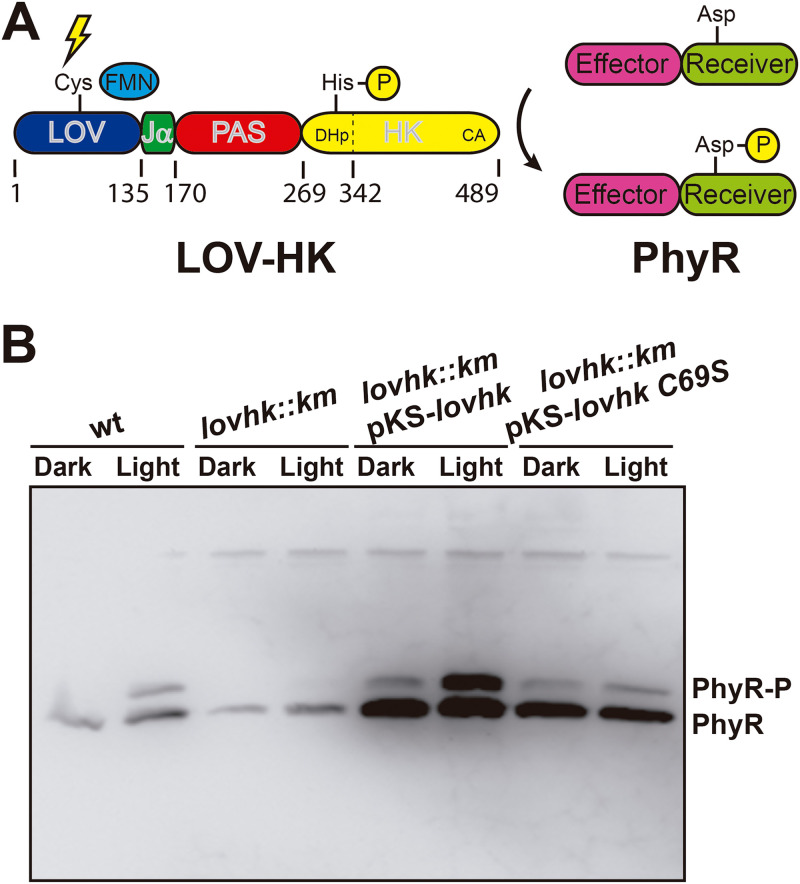
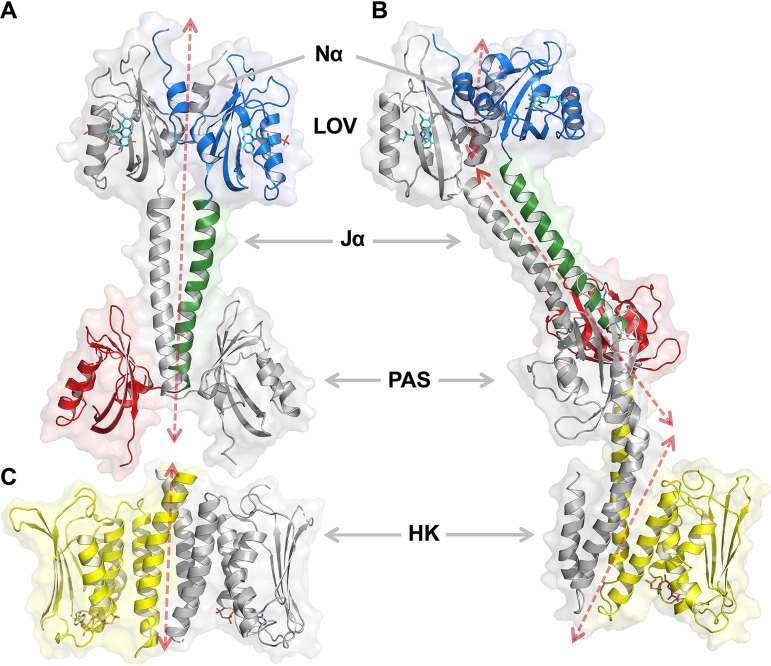
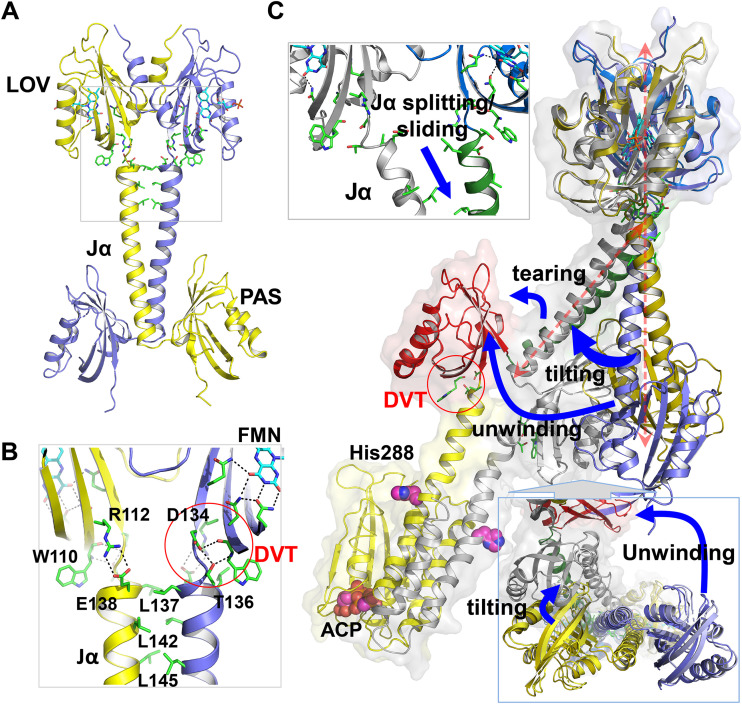
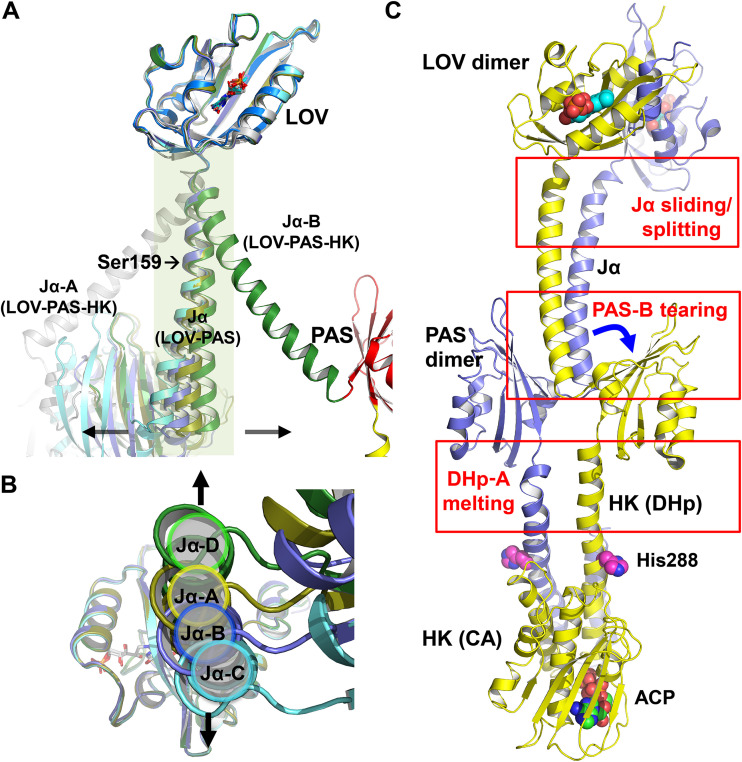
The ability to sense and respond to environmental cues is essential for adaptation and survival in living organisms. In bacteria, this process is accomplished by multidomain sensor histidine kinases that undergo autophosphorylation in response to specific stimuli, thereby triggering downstream signaling cascades. However, the molecular mechanism of allosteric activation is not fully understood in these important sensor proteins. Here, we report the full-length crystal structure of a blue light photoreceptor LOV histidine kinase (LOV-HK) involved in light-dependent virulence modulation in the pathogenic bacterium Brucella abortus Joint analyses of dark and light structures determined in different signaling states have shown that LOV-HK transitions from a symmetric dark structure to a highly asymmetric light state. The initial local and subtle structural signal originated in the chromophore-binding LOV domain alters the dimer asymmetry via a coiled-coil rotary switch and helical bending in the helical spine. These amplified structural changes result in enhanced conformational flexibility and large-scale rearrangements that facilitate the phosphoryl transfer reaction in the HK domain.IMPORTANCE Bacteria employ two-component systems (TCSs) to sense and respond to changes in their surroundings. At the core of the TCS signaling pathway is the multidomain sensor histidine kinase, where the enzymatic activity of its output domain is allosterically controlled by the input signal perceived by the sensor domain. Here, we examine the structures and dynamics of a naturally occurring light-sensitive histidine kinase from the pathogen Brucella abortus in both its full-length and its truncated constructs. Direct comparisons between the structures captured in different signaling states have revealed concerted protein motions in an asymmetric dimer framework in response to light. Findings of this work provide mechanistic insights into modular sensory proteins that share a similar modular architecture.
Keywords: crystallography; dimer asymmetry; light activation mechanism; photoreceptor; sensory histidine kinase.
Copyright © 2021 Rinaldi et al.
Figures
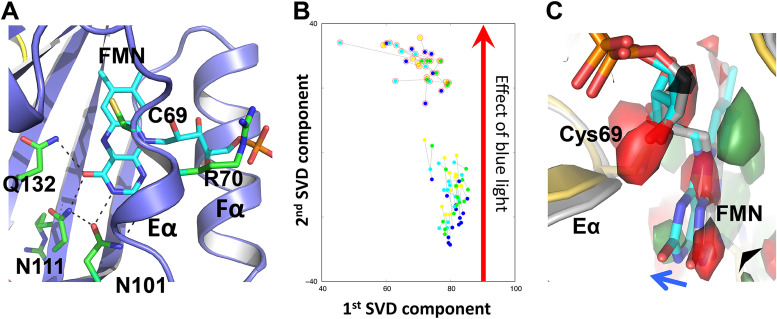
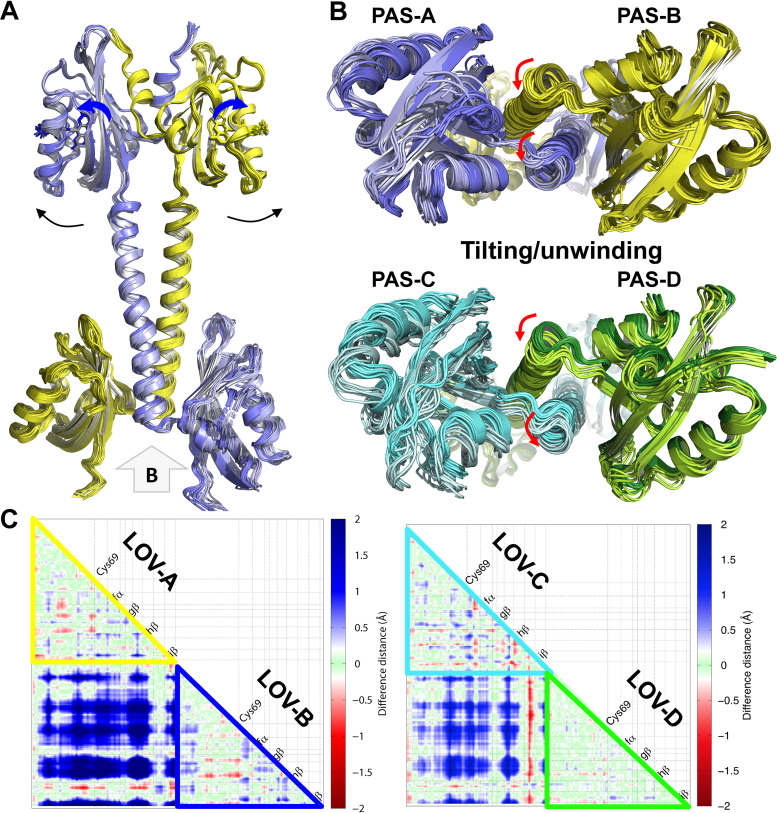
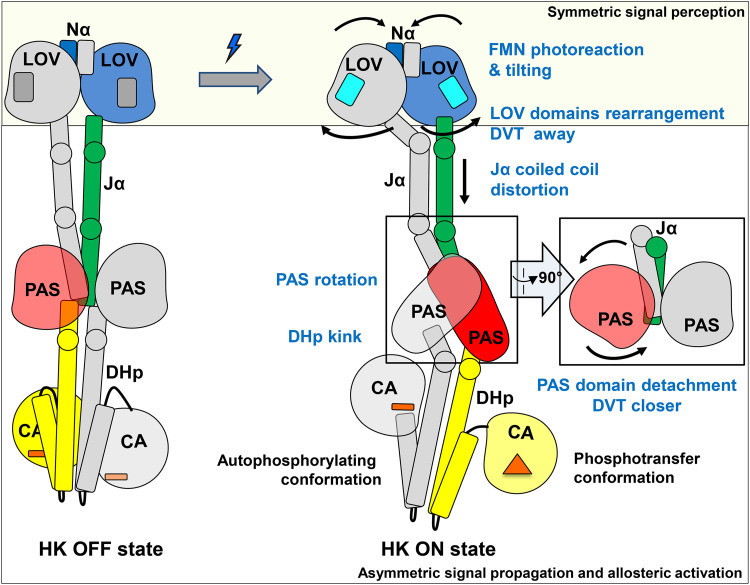
References
-
- Swartz TE, Tseng TS, Frederickson MA, Paris G, Comerci DJ, Rajashekara G, Kim JG, Mudgett MB, Splitter GA, Ugalde RA, Goldbaum FA, Briggs WR, Bogomolni RA. 2007. Blue-light-activated histidine kinases: two-component sensors in bacteria. Science 317:1090–1093. doi:10.1126/science.1144306. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
