

Long-term depression at hippocampal mossy fiber-CA3 synapses involves BDNF but is not mediated by p75NTR signaling
- PMID: 33879805
- PMCID: PMC8058084
- DOI: 10.1038/s41598-021-87769-9
Long-term depression at hippocampal mossy fiber-CA3 synapses involves BDNF but is not mediated by p75NTR signaling
Abstract
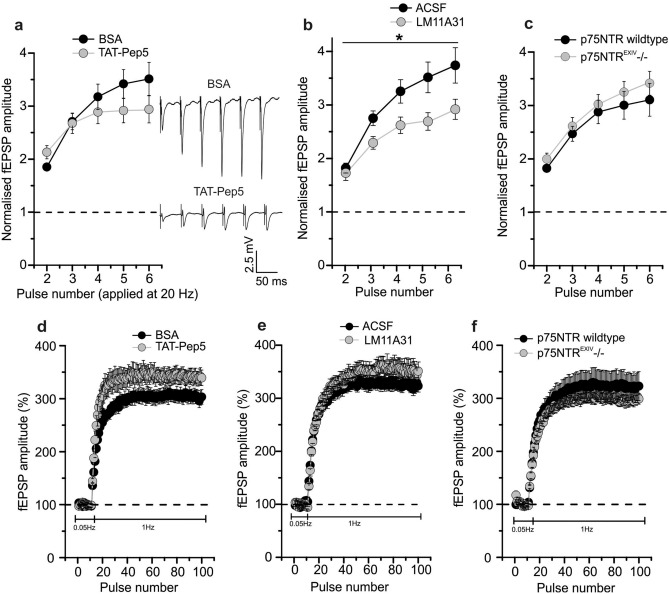
BDNF plays a crucial role in the regulation of synaptic plasticity. It is synthesized as a precursor (proBDNF) that can be proteolytically cleaved to mature BDNF (mBDNF). Previous studies revealed a bidirectional mode of BDNF actions, where long-term potentiation (LTP) was mediated by mBDNF through tropomyosin related kinase (Trk) B receptors whereas long-term depression (LTD) depended on proBDNF/p75 neurotrophin receptor (p75NTR) signaling. While most experimental evidence for this BDNF dependence of synaptic plasticity in the hippocampus was derived from Schaffer collateral (SC)-CA1 synapses, much less is known about the mechanisms of synaptic plasticity, in particular LTD, at hippocampal mossy fiber (MF) synapses onto CA3 neurons. Since proBDNF and mBDNF are expressed most abundantly at MF-CA3 synapses in the rodent brain and we had shown previously that MF-LTP depends on mBDNF/TrkB signaling, we now explored the role of proBDNF/p75NTR signaling in MF-LTD. Our results show that neither acute nor chronic inhibition of p75NTR signaling impairs MF-LTD, while short-term plasticity, in particular paired-pulse facilitation, at MF-CA3 synapses is affected by a lack of functional p75NTR signaling. Furthermore, MF-CA3 synapses showed normal LTD upon acute inhibition of TrkB receptor signaling. Nonetheless, acute inhibition of plasminogen activator inhibitor-1 (PAI-1), an inhibitor of both intracellular and extracellular proBDNF cleavage, impaired MF-LTD. This seems to indicate that LTD at MF-CA3 synapses involves BDNF, however, MF-LTD does not depend on p75NTRs. Altogether, our experiments demonstrate that p75NTR signaling is not warranted for all glutamatergic synapses but rather needs to be checked separately for every synaptic connection.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
