

IFITM3 incorporation sensitizes influenza A virus to antibody-mediated neutralization
- PMID: 33882122
- PMCID: PMC8072448
- DOI: 10.1084/jem.20200303
IFITM3 incorporation sensitizes influenza A virus to antibody-mediated neutralization
Abstract
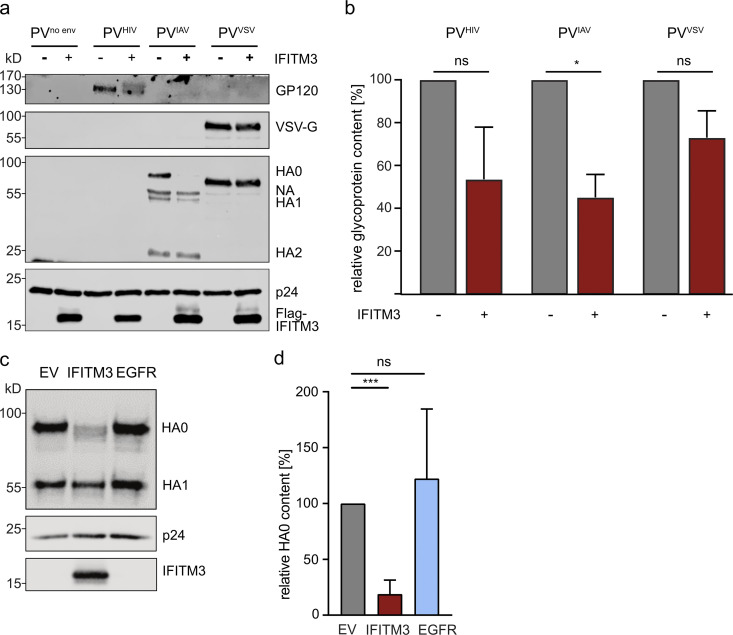
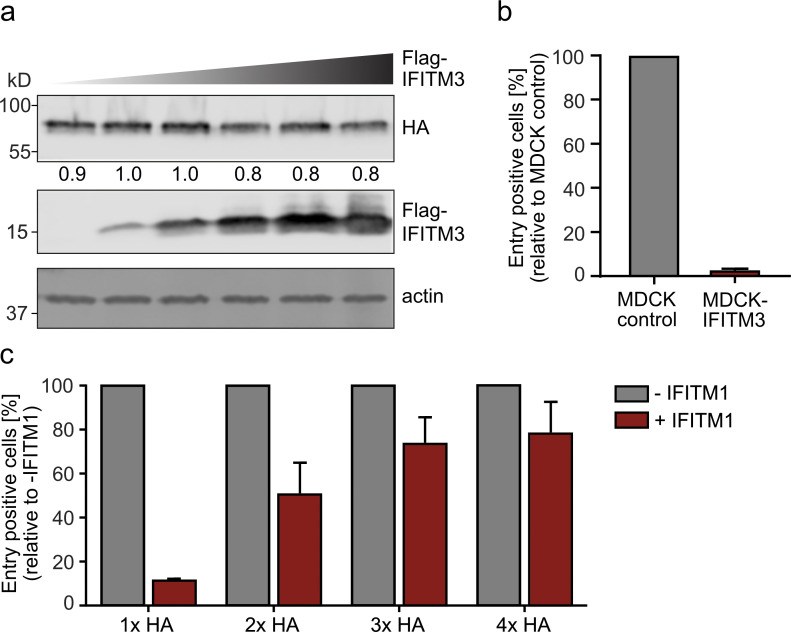
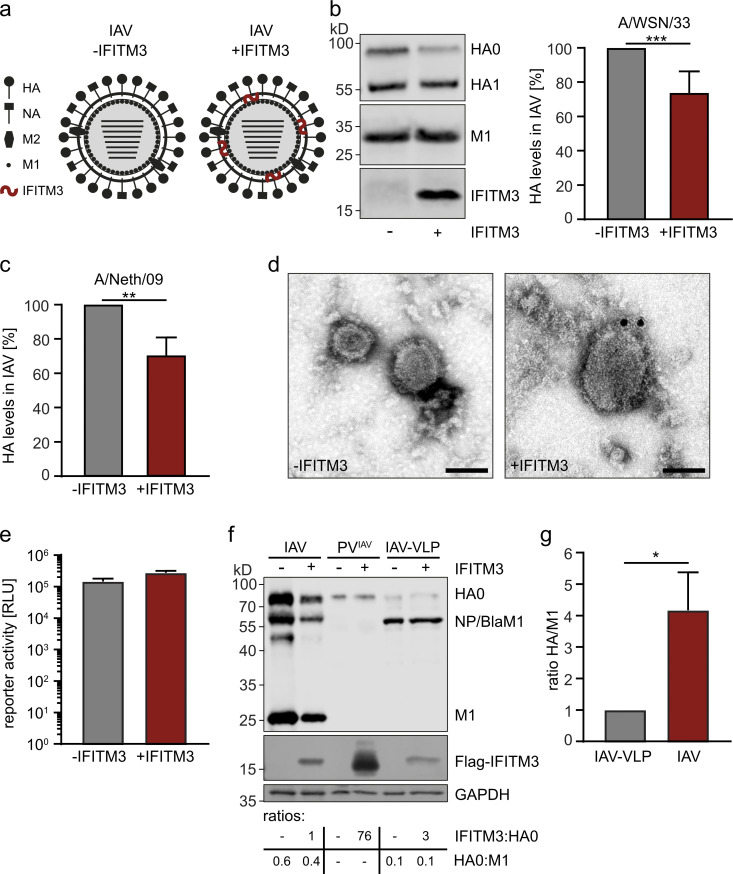
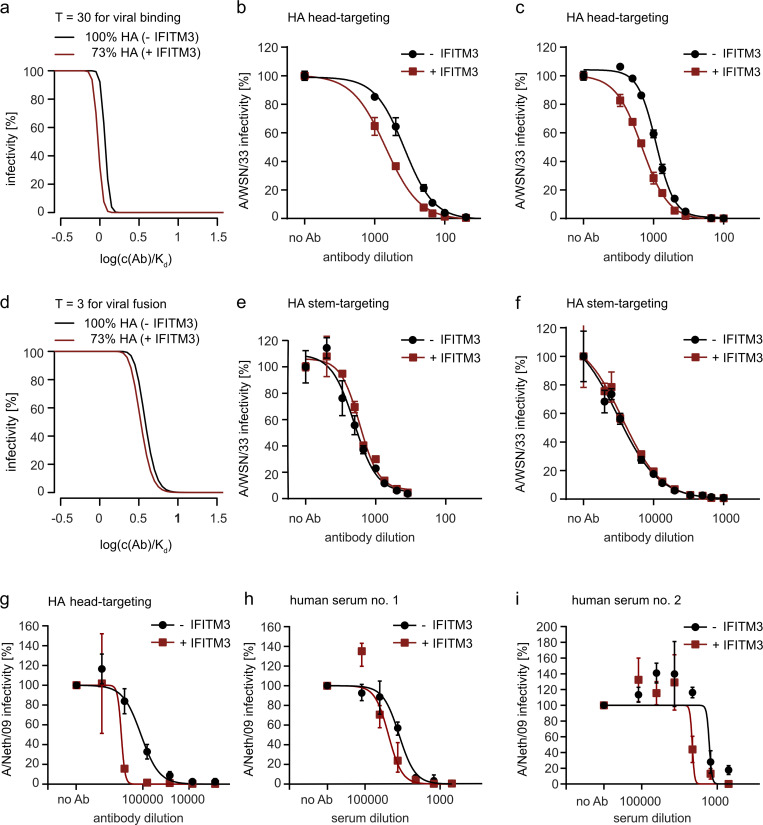
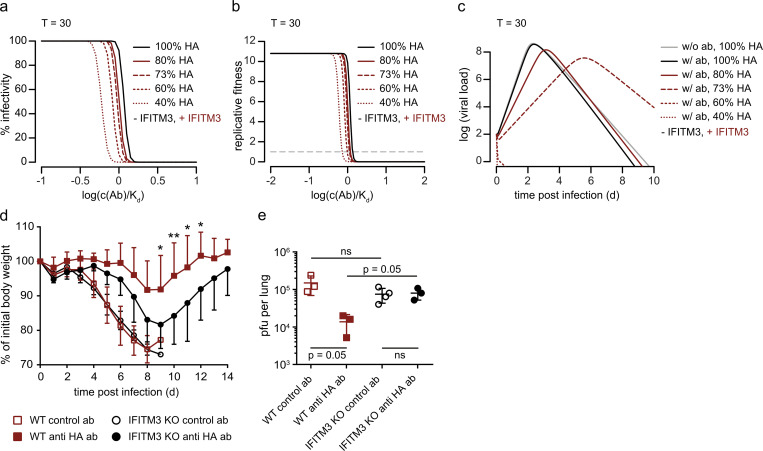
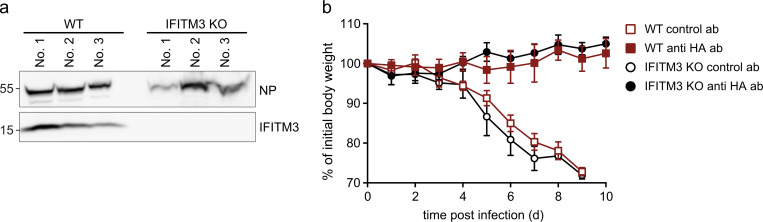
The disease severity of influenza is highly variable in humans, and one genetic determinant behind these differences is the IFITM3 gene. As an effector of the interferon response, IFITM3 potently blocks cytosolic entry of influenza A virus (IAV). Here, we reveal a novel level of inhibition by IFITM3 in vivo: We show that incorporation of IFITM3 into IAV particles competes with incorporation of viral hemagglutinin (HA). Decreased virion HA levels did not reduce infectivity, suggesting that high HA density on IAV virions may be an antagonistic strategy used by the virus to prevent direct inhibition. However, we found that IFITM3-mediated reduction in HA content sensitizes IAV to antibody-mediated neutralization. Mathematical modeling predicted that this effect decreases and delays peak IAV titers, and we show that, indeed, IFITM3-mediated sensitization of IAV to antibody-mediated neutralization impacts infection outcome in an in vivo mouse model. Overall, our data describe a previously unappreciated interplay between the innate effector IFITM3 and the adaptive immune response.
© 2021 Lanz et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures
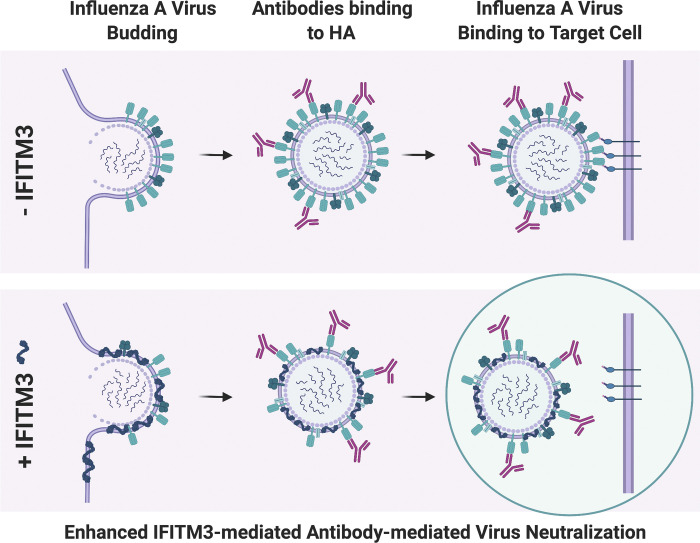
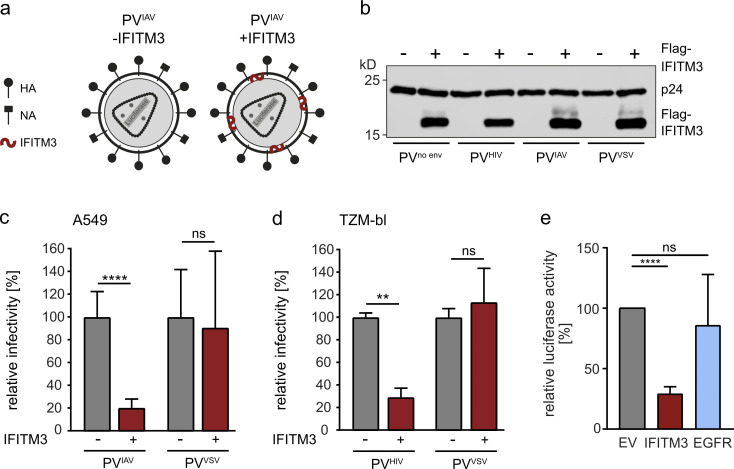
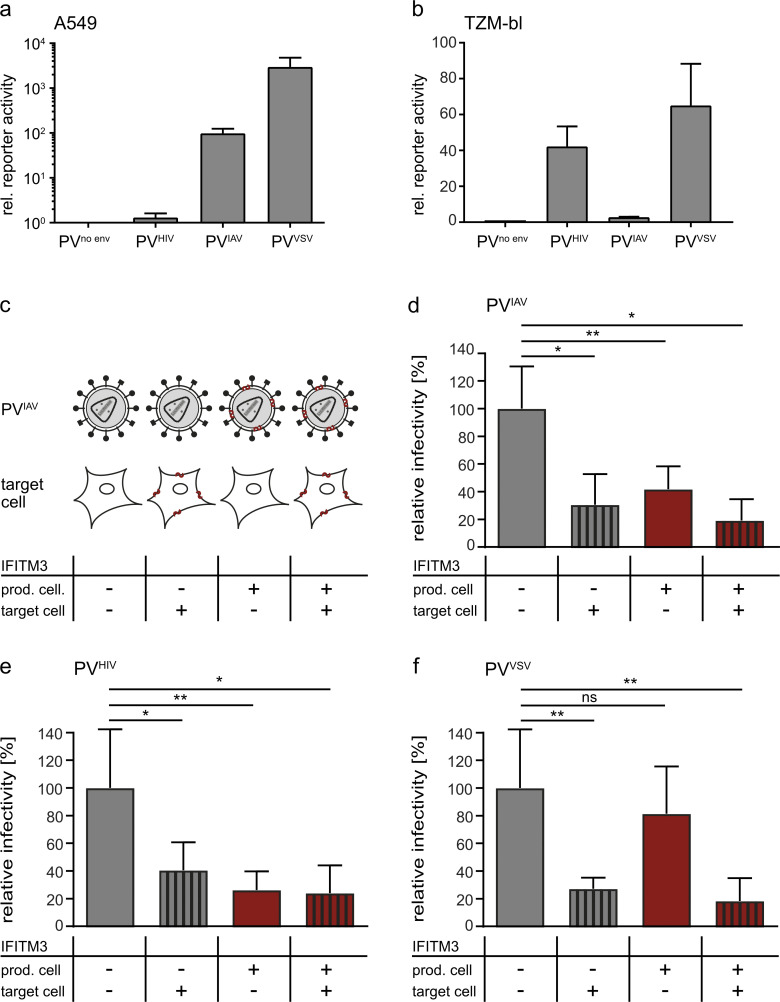
References
-
- Allen, E.K., Randolph A.G., Bhangale T., Dogra P., Ohlson M., Oshansky C.M., Zamora A.E., Shannon J.P., Finkelstein D., Dressen A., et al. . 2017. SNP-mediated disruption of CTCF binding at the IFITM3 promoter is associated with risk of severe influenza in humans. Nat. Med. 23:975–983. 10.1038/nm.4370 - DOI - PMC - PubMed
-
- Appourchaux, R., Delpeuch M., Zhong L., Burlaud-Gaillard J., Tartour K., Savidis G., Brass A., Etienne L., Roingeard P., and Cimarelli A.. 2019. Functional Mapping of Regions Involved in the Negative Imprinting of Virion Particle Infectivity and in Target Cell Protection by Interferon-Induced Transmembrane Protein 3 against HIV-1. J. Virol. 93:e01716-18. 10.1128/JVI.01716-18 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
