

Restoration of visual function in advanced disease after transplantation of purified human pluripotent stem cell-derived cone photoreceptors
- PMID: 33882303
- PMCID: PMC8065177
- DOI: 10.1016/j.celrep.2021.109022
Restoration of visual function in advanced disease after transplantation of purified human pluripotent stem cell-derived cone photoreceptors
Abstract
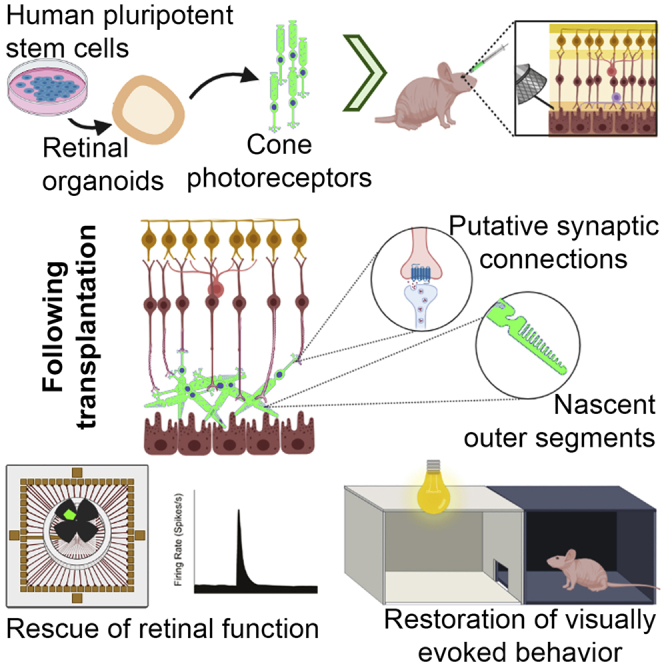
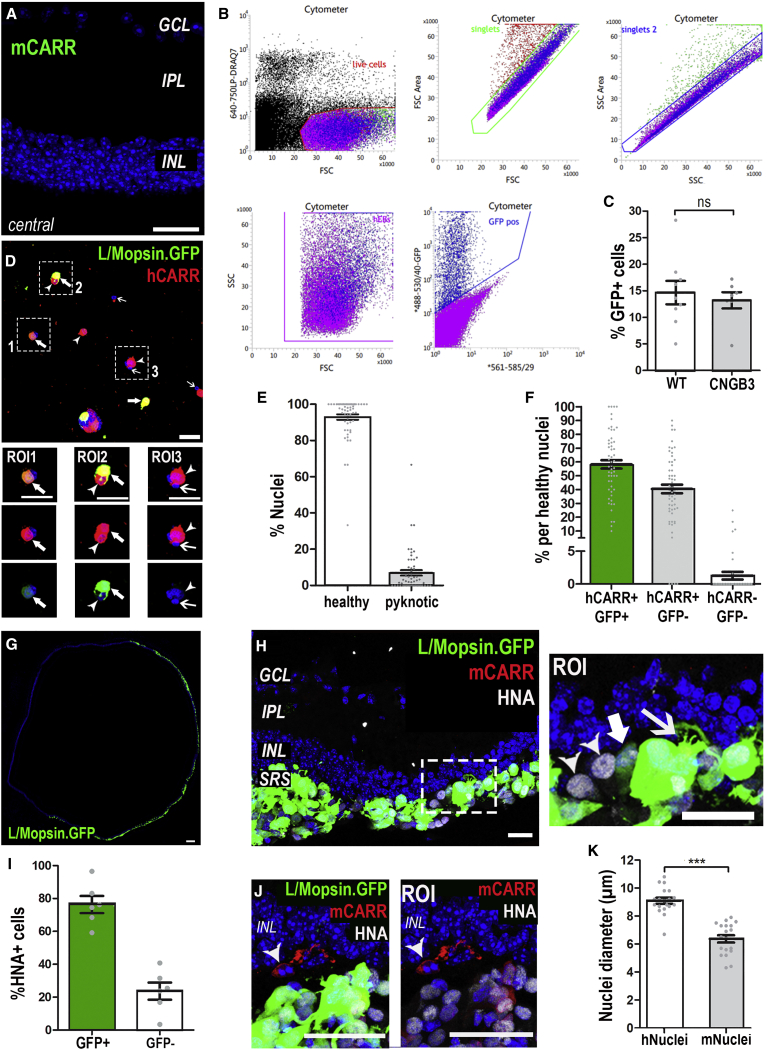
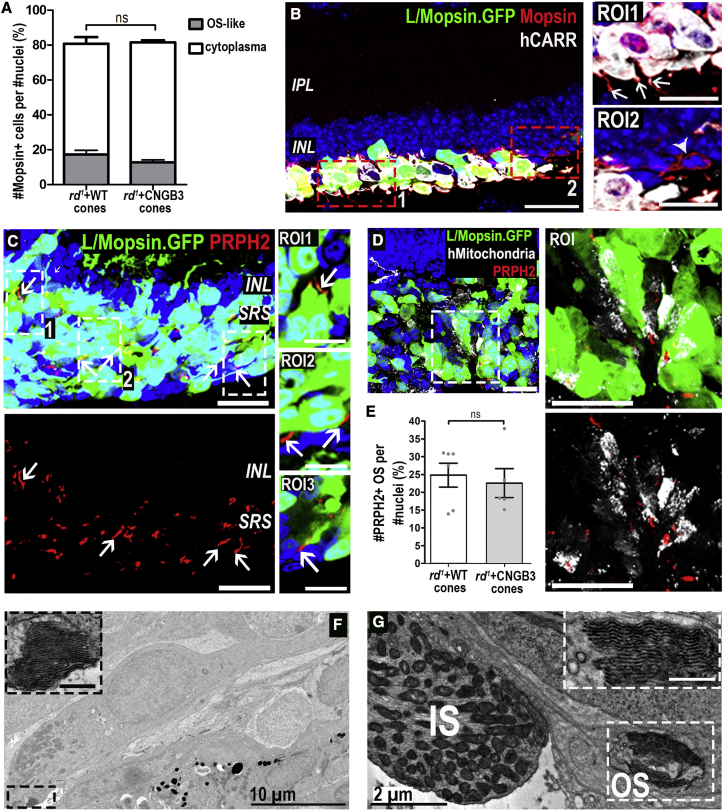
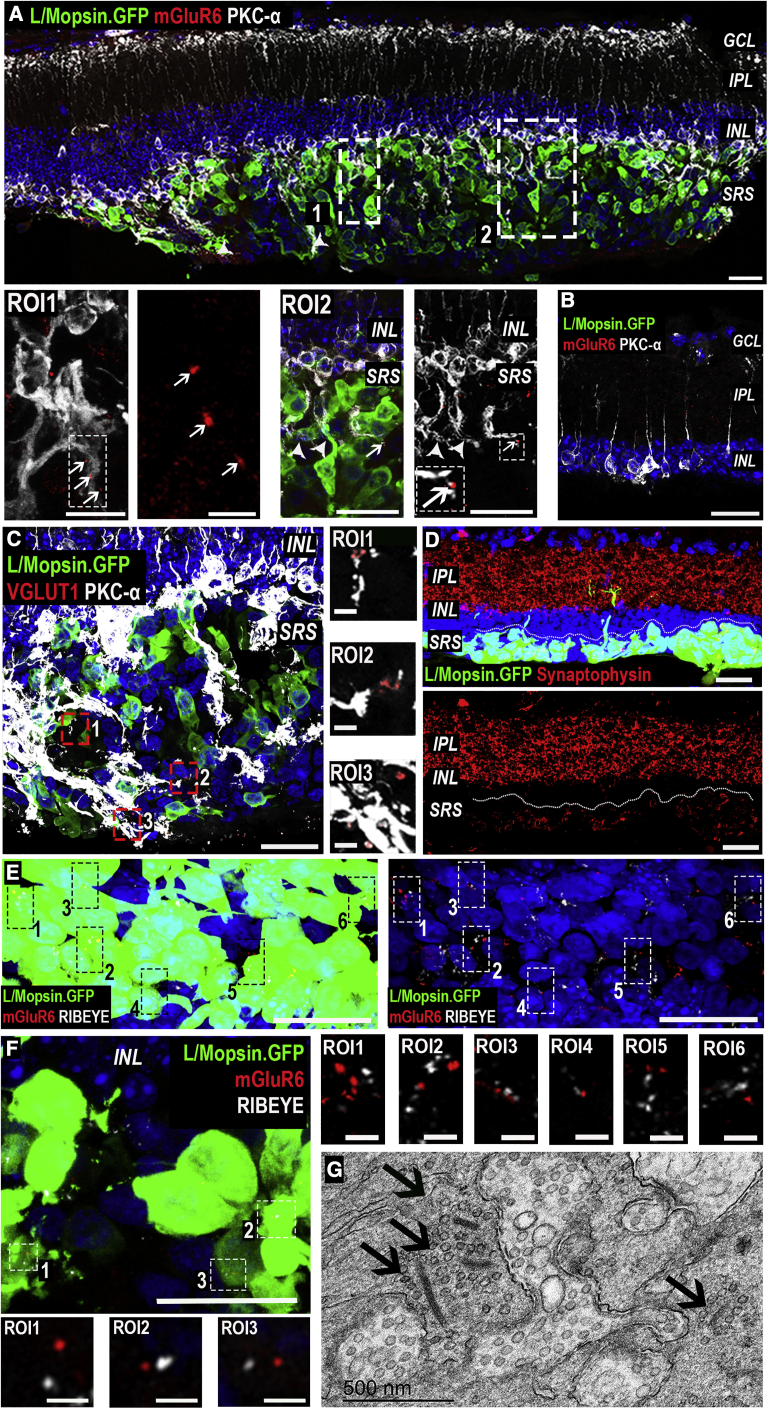
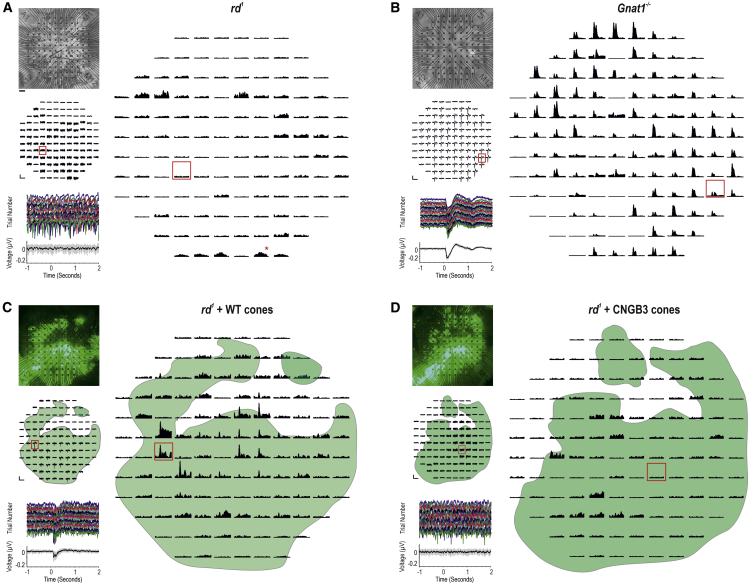
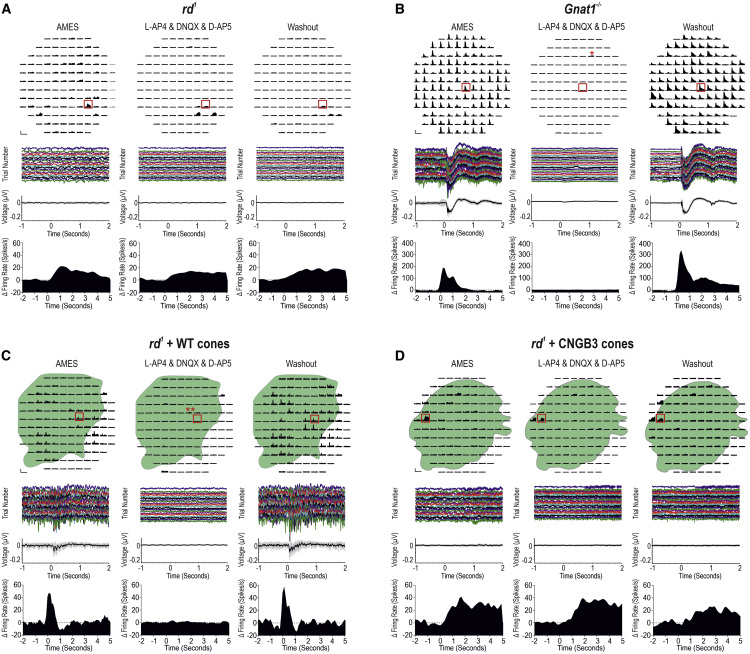
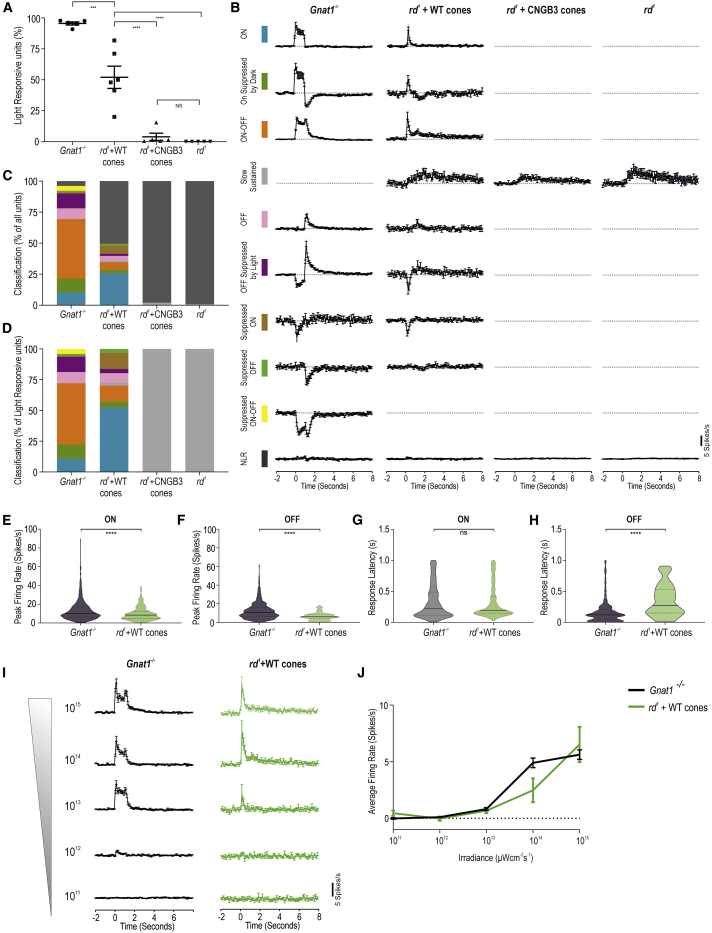
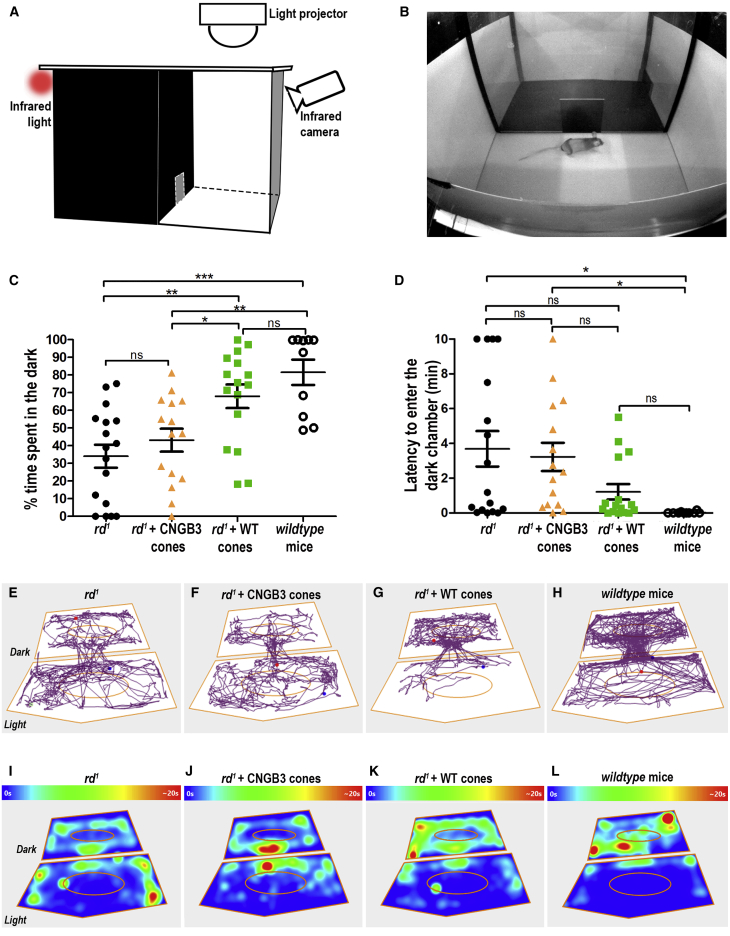
Age-related macular degeneration and other macular diseases result in the loss of light-sensing cone photoreceptors, causing irreversible sight impairment. Photoreceptor replacement may restore vision by transplanting healthy cells, which must form new synaptic connections with the recipient retina. Despite recent advances, convincing evidence of functional connectivity arising from transplanted human cone photoreceptors in advanced retinal degeneration is lacking. Here, we show restoration of visual function after transplantation of purified human pluripotent stem cell-derived cones into a mouse model of advanced degeneration. Transplanted human cones elaborate nascent outer segments and make putative synapses with recipient murine bipolar cells (BCs), which themselves undergo significant remodeling. Electrophysiological and behavioral assessments demonstrate restoration of surprisingly complex light-evoked retinal ganglion cell responses and improved light-evoked behaviors in treated animals. Stringent controls exclude alternative explanations, including material transfer and neuroprotection. These data provide crucial validation for photoreceptor replacement therapy and for the potential to rescue cone-mediated vision.
Keywords: cell therapy; cone photoreceptor; degeneration; electrophysiology; macular degeneration; outer segment; rescue; retinal organoid; transplantation; visual function.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Aghaizu N.D., Kruczek K., Gonzalez-Cordero A., Ali R.R., Pearson R.A. Pluripotent stem cells and their utility in treating photoreceptor degenerations. Prog. Brain Res. 2017;231:191–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
