

Subiculum as a generator of sharp wave-ripples in the rodent hippocampus
- PMID: 33882307
- PMCID: PMC9239734
- DOI: 10.1016/j.celrep.2021.109021
Subiculum as a generator of sharp wave-ripples in the rodent hippocampus
Abstract
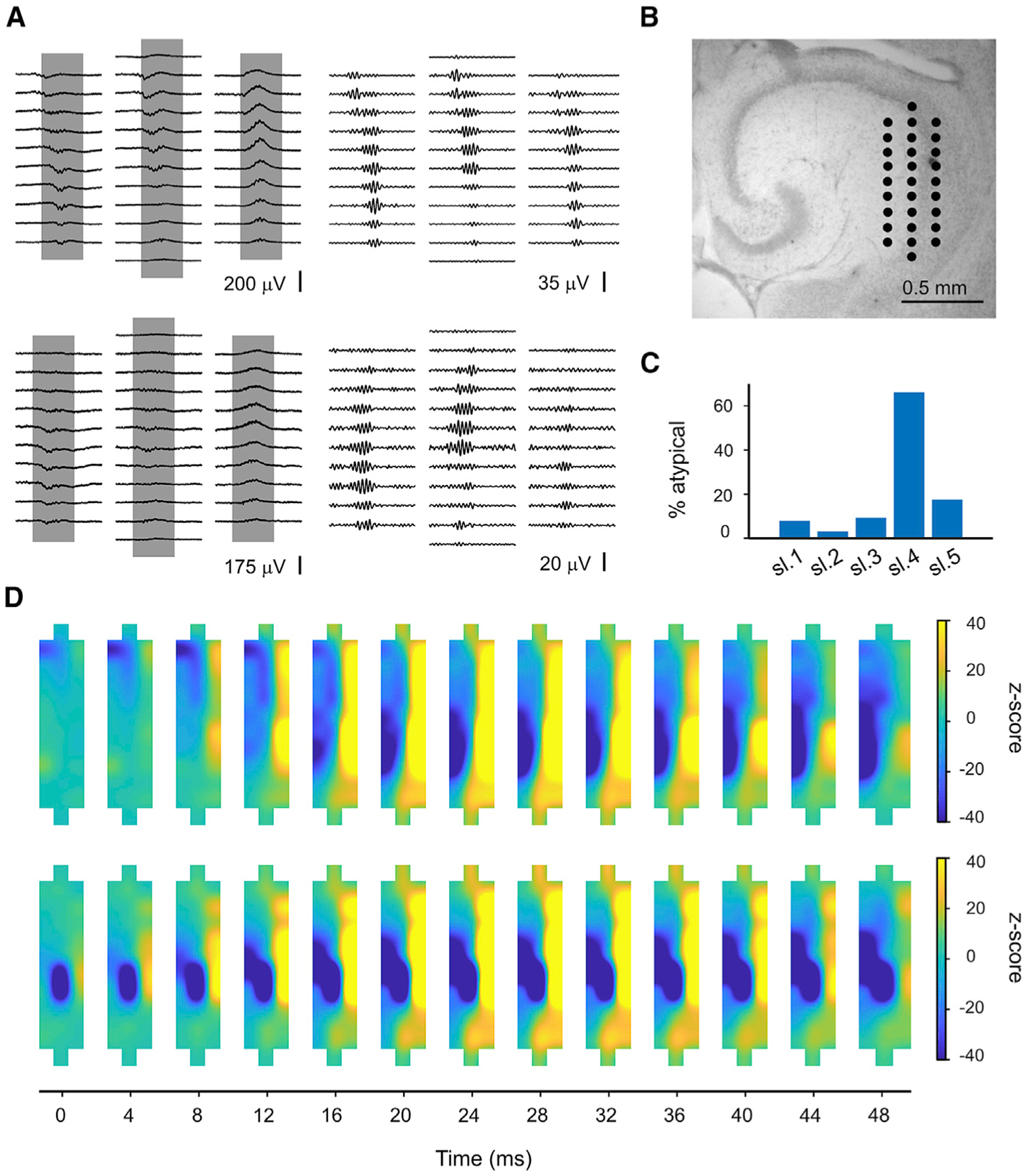
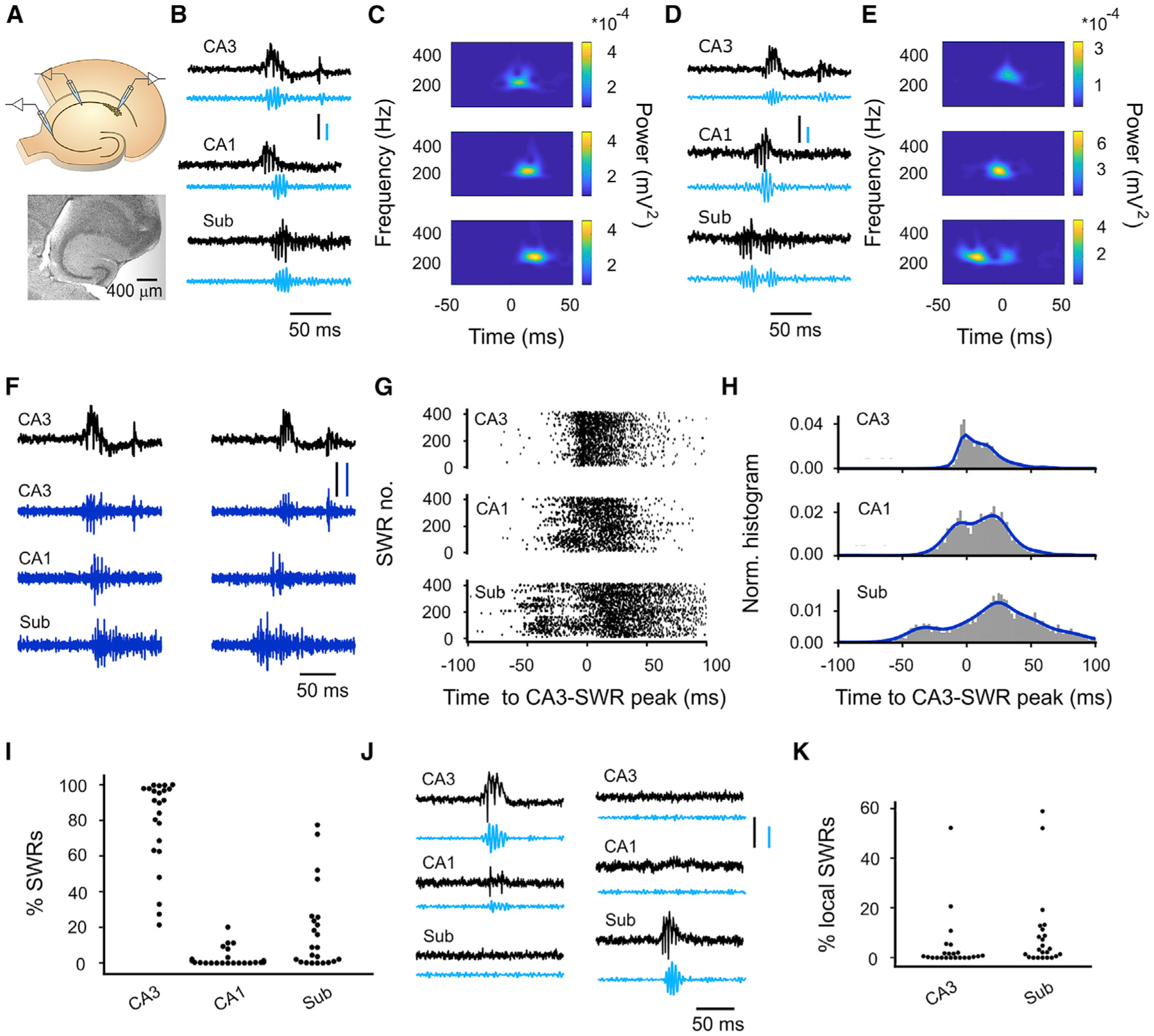
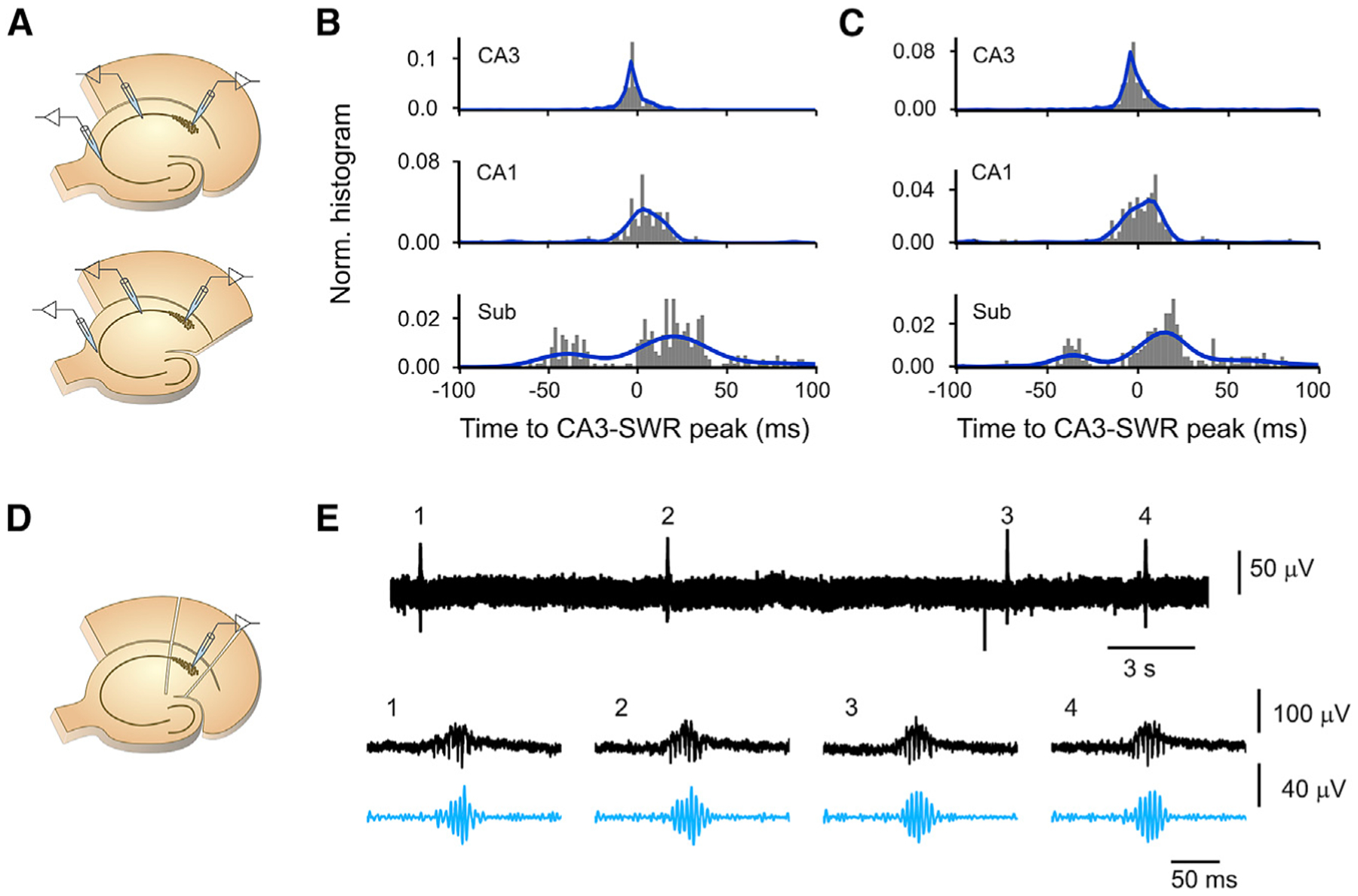
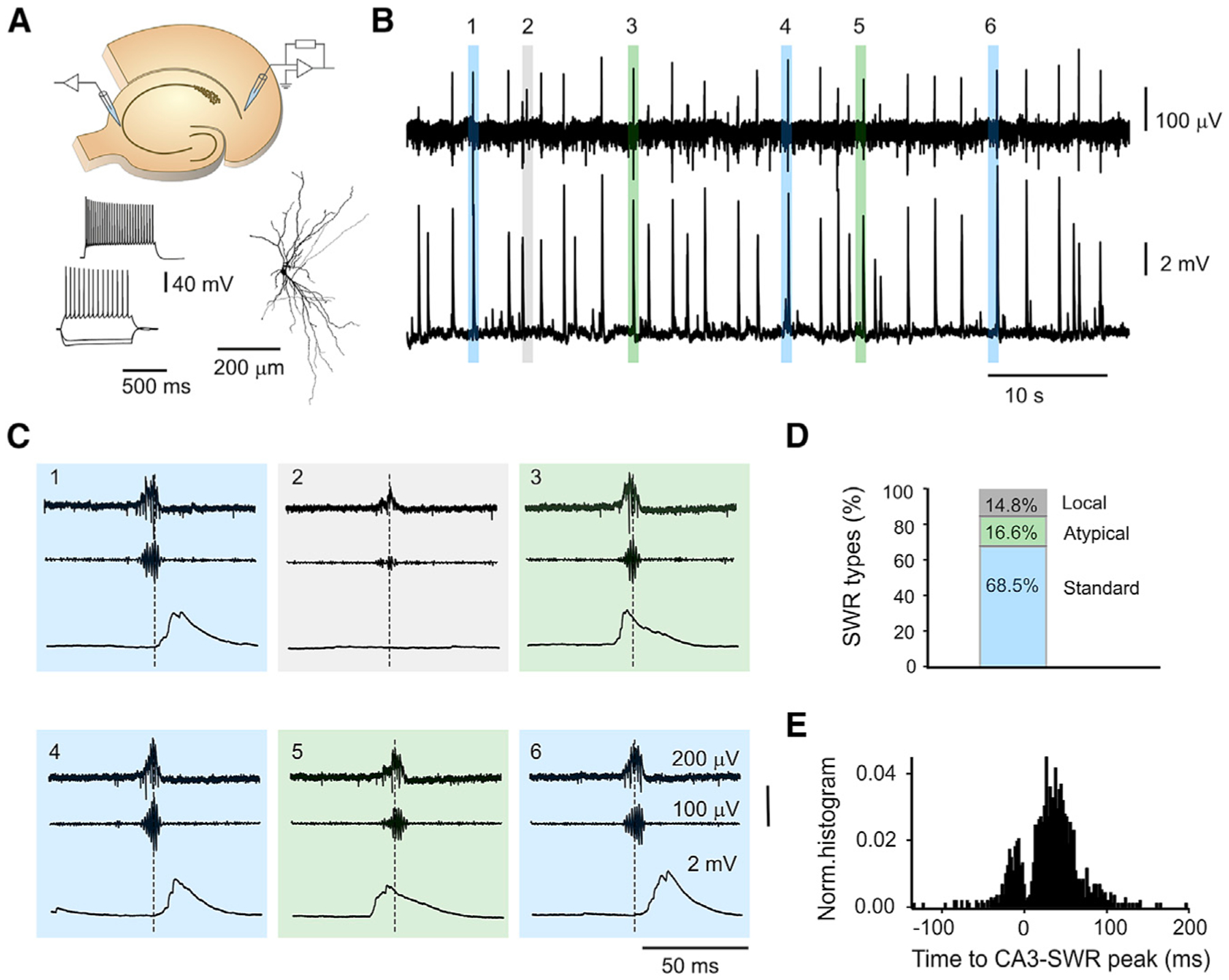
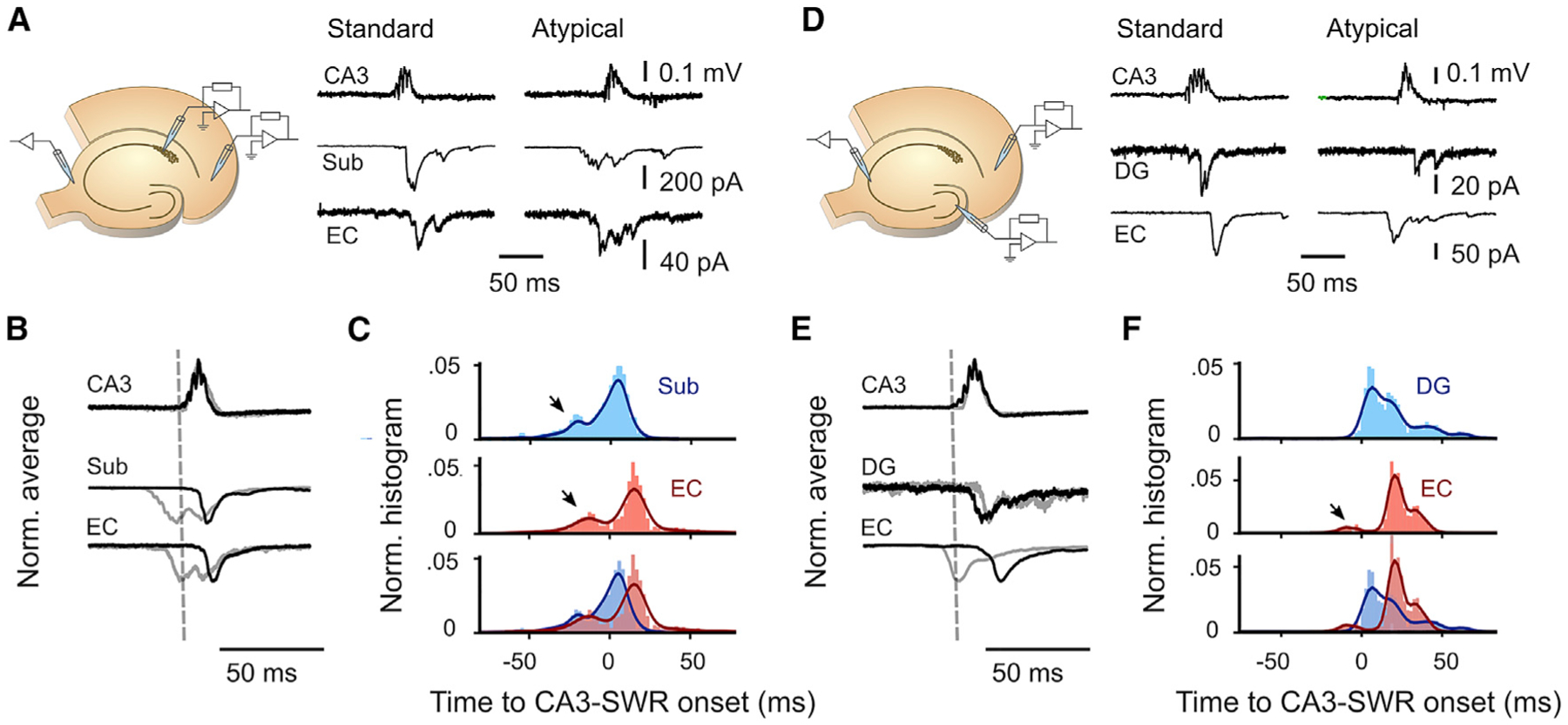
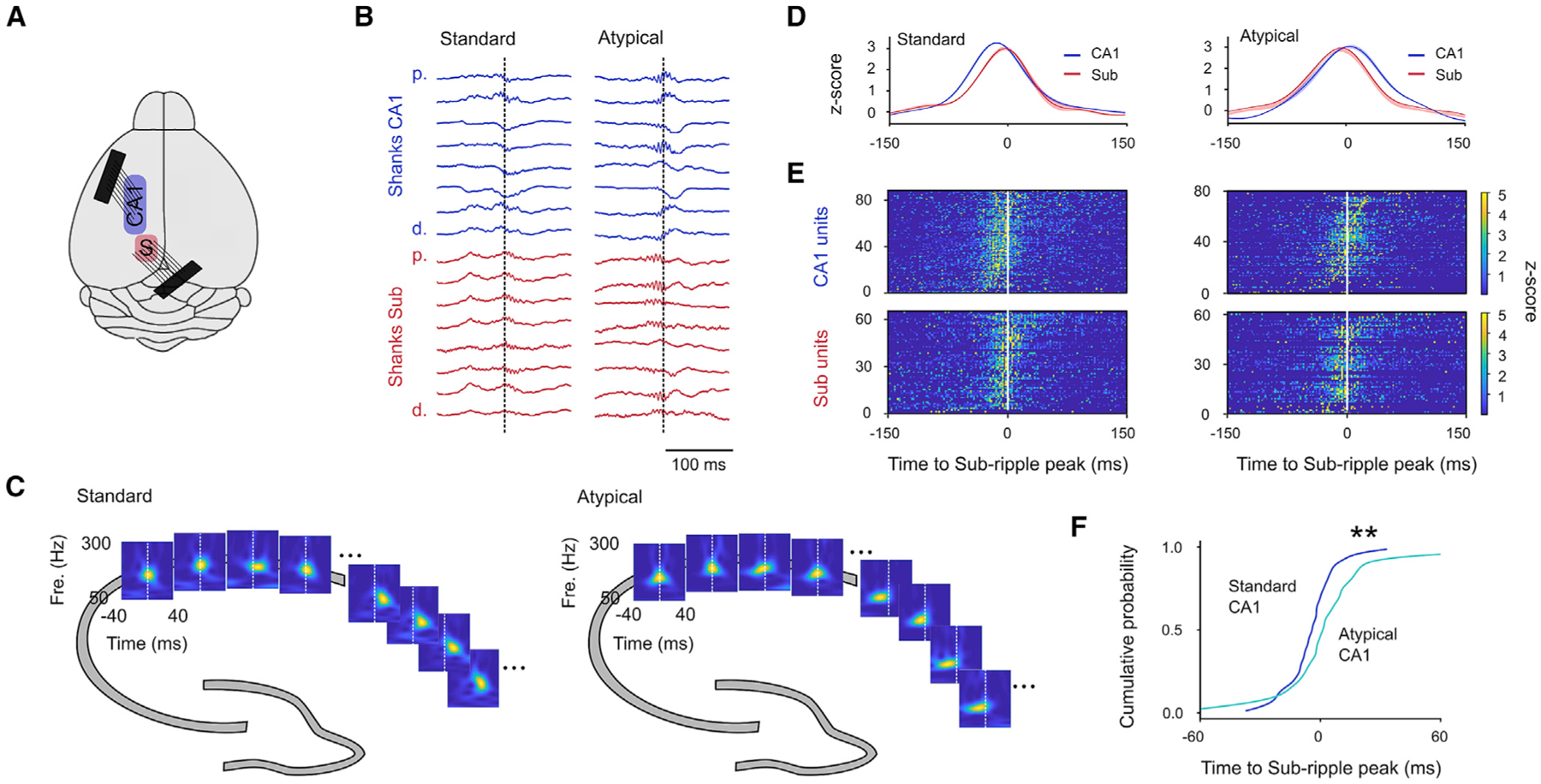
Sharp wave-ripples (SWRs) represent synchronous discharges of hippocampal neurons and are believed to play a major role in memory consolidation. A large body of evidence suggests that SWRs are exclusively generated in the CA3-CA2 network. In contrast, here, we provide several lines of evidence showing that the subiculum can function as a secondary SWRs generator. SWRs with subicular origin propagate forward into the entorhinal cortex as well as backward into the hippocampus proper. Our findings suggest that the output structures of the hippocampus are not only passively facilitating the transfer of SWRs to the cortex, but they also can actively contribute to the genesis of SWRs. We hypothesize that SWRs with a subicular origin may be important for the consolidation of information conveyed to the hippocampus via the temporoammonic pathway.
Keywords: CA1; CA3; entorhinal cortex; hippocampus; oscillations; sharp wave-ripples; subiculum.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Downstream effects of hippocampal sharp wave ripple oscillations on medial entorhinal cortex layer V neurons in vitro.Hippocampus. 2016 Dec;26(12):1493-1508. doi: 10.1002/hipo.22623. Epub 2016 Aug 18. Hippocampus. 2016. PMID: 27479916
-
Circuit mechanisms of hippocampal reactivation during sleep.Neurobiol Learn Mem. 2019 Apr;160:98-107. doi: 10.1016/j.nlm.2018.04.018. Epub 2018 May 1. Neurobiol Learn Mem. 2019. PMID: 29723670 Free PMC article.
-
Subicular activation preceding hippocampal ripples in vitro.Sci Rep. 2013;3:2696. doi: 10.1038/srep02696. Sci Rep. 2013. PMID: 24045268 Free PMC article.
-
Progress on the hippocampal circuits and functions based on sharp wave ripples.Brain Res Bull. 2023 Aug;200:110695. doi: 10.1016/j.brainresbull.2023.110695. Epub 2023 Jun 21. Brain Res Bull. 2023. PMID: 37353037 Review.
-
CA2 orchestrates hippocampal network dynamics.Hippocampus. 2023 Mar;33(3):241-251. doi: 10.1002/hipo.23495. Epub 2022 Dec 27. Hippocampus. 2023. PMID: 36575880 Free PMC article. Review.
Cited by
-
Advanced age has dissociable effects on hippocampal CA1 ripples and CA3 high frequency events in male rats.Neurobiol Aging. 2022 Sep;117:44-58. doi: 10.1016/j.neurobiolaging.2022.04.014. Epub 2022 May 5. Neurobiol Aging. 2022. PMID: 35665647 Free PMC article.
-
Rhythmic Memory Consolidation in the Hippocampus.Front Neural Circuits. 2022 Apr 1;16:885684. doi: 10.3389/fncir.2022.885684. eCollection 2022. Front Neural Circuits. 2022. PMID: 35431819 Free PMC article.
-
Differential ripple propagation along the hippocampal longitudinal axis.Elife. 2023 Apr 13;12:e85488. doi: 10.7554/eLife.85488. Elife. 2023. PMID: 37052307 Free PMC article.
-
Subicular neurons represent multiple variables of a hippocampal-dependent task by using theta rhythm.PLoS Biol. 2022 Jan 31;20(1):e3001546. doi: 10.1371/journal.pbio.3001546. eCollection 2022 Jan. PLoS Biol. 2022. PMID: 35100261 Free PMC article.
-
Cell-specific switch for epileptiform activity: critical role of interneurons in the mouse subicular network.Cereb Cortex. 2023 May 9;33(10):6171-6183. doi: 10.1093/cercor/bhac493. Cereb Cortex. 2023. PMID: 36611229 Free PMC article.
References
-
- Bragin A, Jandó G, Nádasdy Z, van Landeghem M, and Buzsáki G (1995). Dentate EEG spikes and associated interneuronal population bursts in the hippocampal hilar region of the rat. J. Neurophysiol 73, 1691–1705. - PubMed
-
- Buzsáki G (1986). Hippocampal sharp waves: their origin and significance. Brain Res. 398, 242–252. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
