

EGFR-dependent tyrosine phosphorylation of integrin β4 is not required for downstream signaling events in cancer cell lines
- PMID: 33883672
- PMCID: PMC8060419
- DOI: 10.1038/s41598-021-88134-6
EGFR-dependent tyrosine phosphorylation of integrin β4 is not required for downstream signaling events in cancer cell lines
Abstract
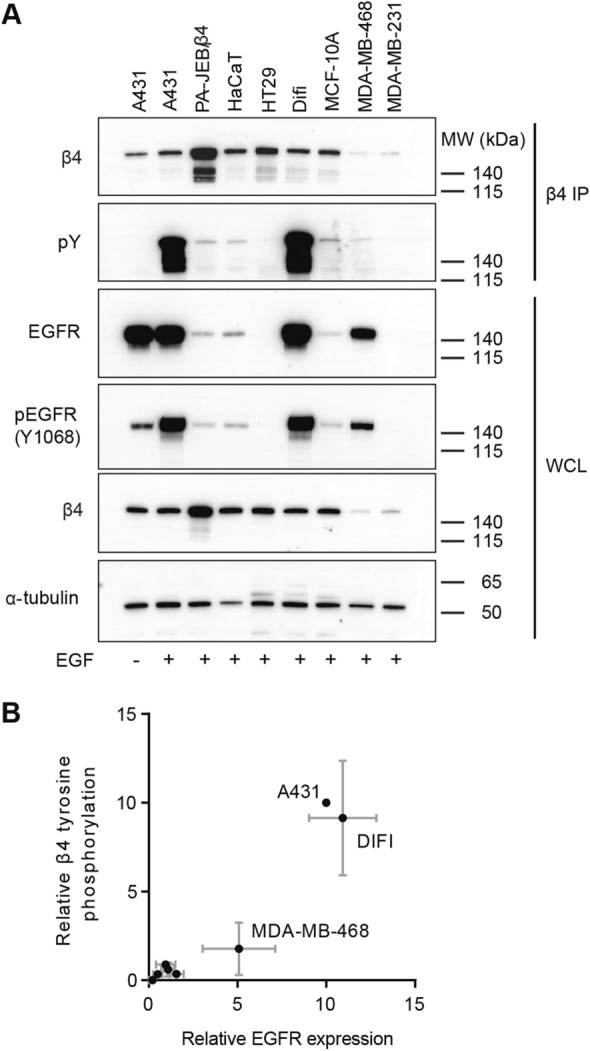
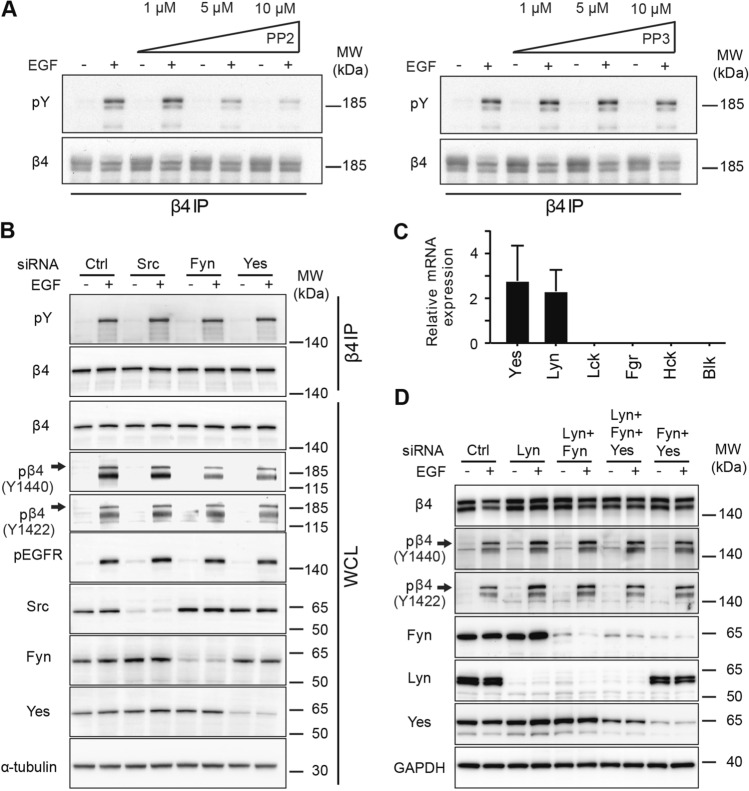
In epithelial cancers, the epidermal growth factor receptor (EGFR) and integrin α6β4 are frequently overexpressed and found to synergistically activate intracellular signaling pathways that promote cell proliferation and migration. In cancer cells, the β4 subunit is phosphorylated at tyrosine residues not normally recognized as kinase substrates; however, the function of these phosphotyrosine residues in cancer cells is a subject of much debate. In EGFR-overexpressing carcinoma cells, we found that the Src family kinase (SFK) inhibitor PP2 reduces β4 tyrosine phosphorylation following the activation of EGFR. However, siRNA mediated knockdown of the SFKs Src, Fyn, Yes and Lyn, individually or in combination, did not affect the EGF-induced phosphorylation of β4. Using phospho-peptide affinity chromatography and mass spectrometry, we found that PLCγ1 binds β4 at the phosphorylated residues Y1422/Y1440, but were unable to verify this interaction in A431 carcinoma cells that overexpress the EGFR. Furthermore, using A431 cells devoid of β4 or reconstituted with phenylalanine specific mutants of β4, the activation of several downstream signaling pathways, including PLCγ/PKC, MAPK and PI3K/Akt, were not substantially affected. We conclude that tyrosine-phosphorylated β4 does not enhance EGFR-mediated signaling in EGFR-overexpressing cells, despite the fact that this integrin subunit is highly tyrosine phosphorylated in these cells.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
