

Selfish chromosomal drive shapes recent centromeric histone evolution in monkeyflowers
- PMID: 33886547
- PMCID: PMC8061799
- DOI: 10.1371/journal.pgen.1009418
Selfish chromosomal drive shapes recent centromeric histone evolution in monkeyflowers
Abstract
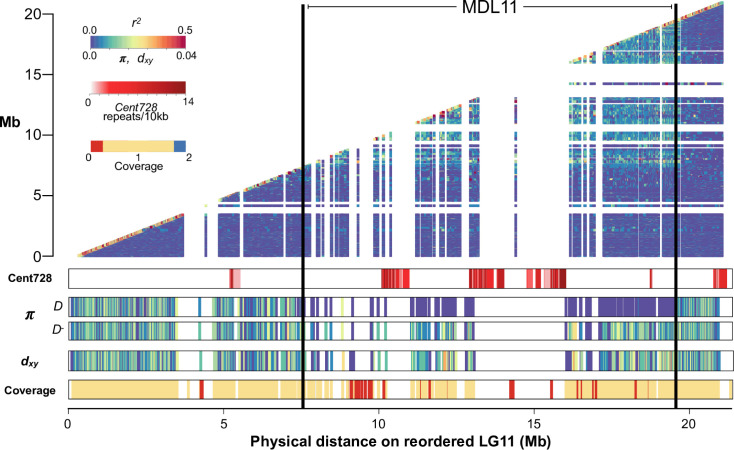
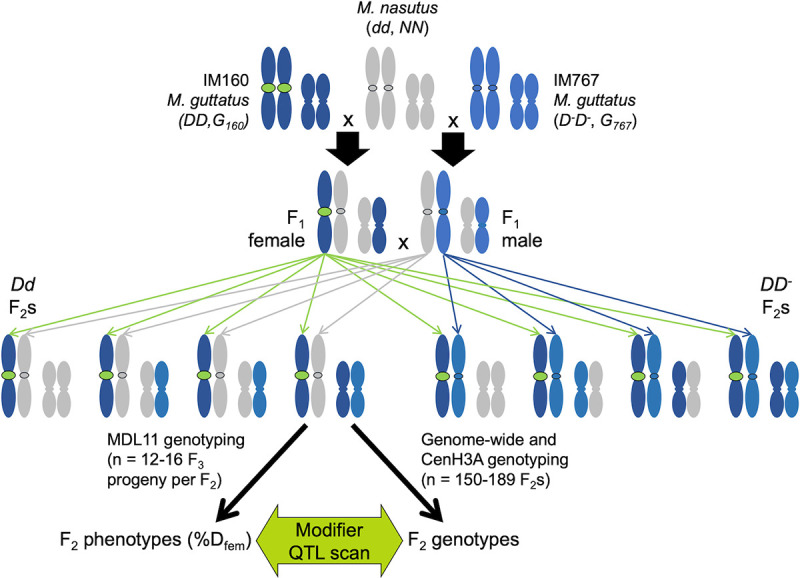
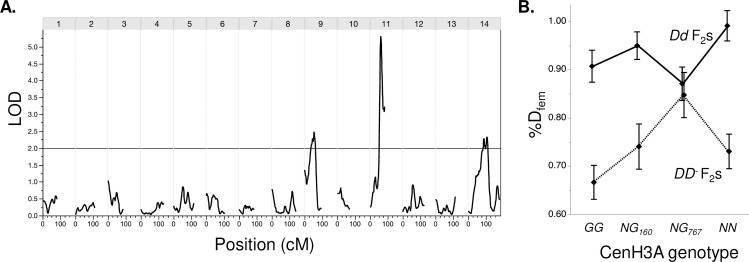
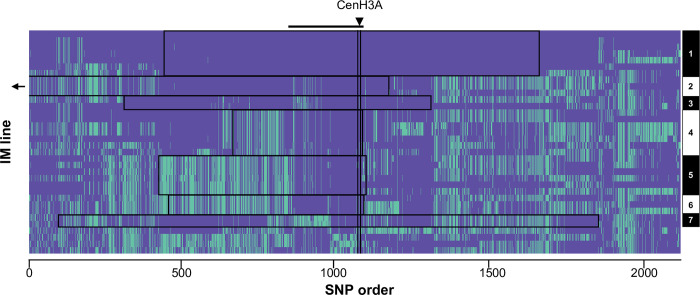
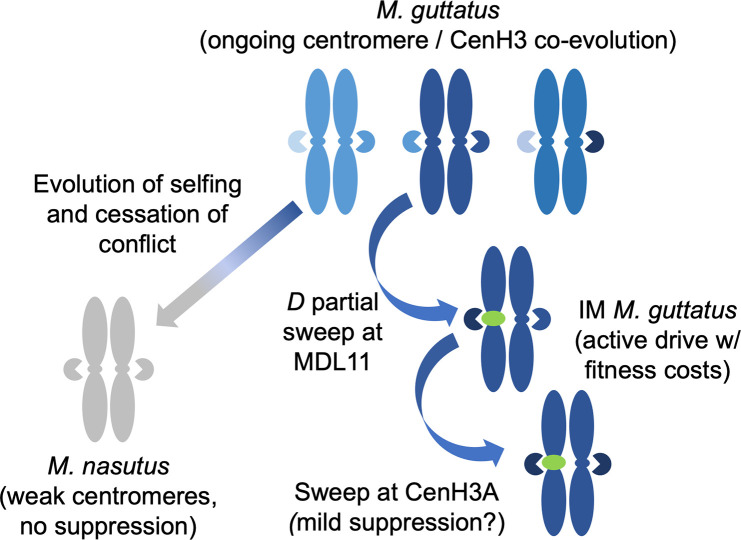
Centromeres are essential mediators of chromosomal segregation, but both centromeric DNA sequences and associated kinetochore proteins are paradoxically diverse across species. The selfish centromere model explains rapid evolution by both components via an arms-race scenario: centromeric DNA variants drive by distorting chromosomal transmission in female meiosis and attendant fitness costs select on interacting proteins to restore Mendelian inheritance. Although it is clear than centromeres can drive and that drive often carries costs, female meiotic drive has not been directly linked to selection on kinetochore proteins in any natural system. Here, we test the selfish model of centromere evolution in a yellow monkeyflower (Mimulus guttatus) population polymorphic for a costly driving centromere (D). We show that the D haplotype is structurally and genetically distinct and swept to a high stable frequency within the past 1500 years. We use quantitative genetic mapping to demonstrate that context-dependence in the strength of drive (from near-100% D transmission in interspecific hybrids to near-Mendelian in within-population crosses) primarily reflects variable vulnerability of the non-driving competitor chromosomes, but also map an unlinked modifier of drive coincident with kinetochore protein Centromere-specific Histone 3 A (CenH3A). Finally, CenH3A exhibits a recent (<1000 years) selective sweep in our focal population, implicating local interactions with D in ongoing adaptive evolution of this kinetochore protein. Together, our results demonstrate an active co-evolutionary arms race between DNA and protein components of the meiotic machinery in Mimulus, with important consequences for individual fitness and molecular divergence.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Putting the brakes on centromere drive in Mimulus.PLoS Genet. 2021 Apr 22;17(4):e1009494. doi: 10.1371/journal.pgen.1009494. eCollection 2021 Apr. PLoS Genet. 2021. PMID: 33886542 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
