

Fast skeletal myosin-binding protein-C regulates fast skeletal muscle contraction
- PMID: 33888578
- PMCID: PMC8092462
- DOI: 10.1073/pnas.2003596118
Fast skeletal myosin-binding protein-C regulates fast skeletal muscle contraction
Abstract
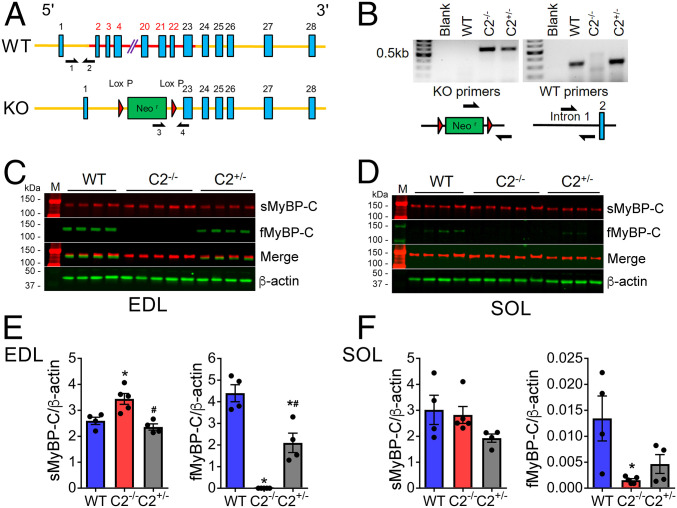
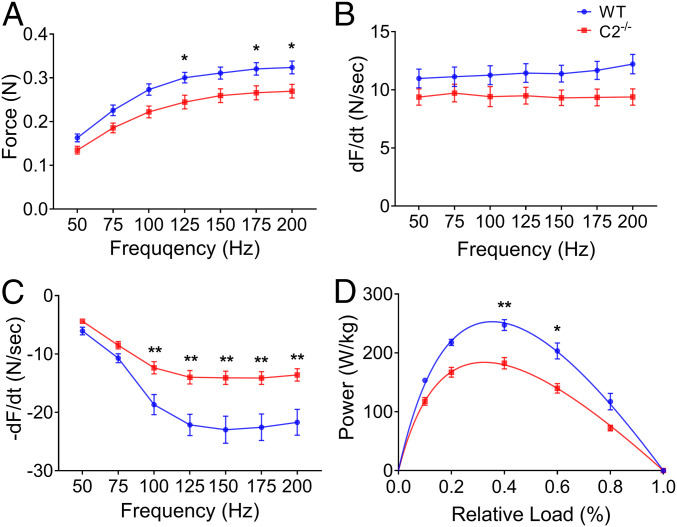
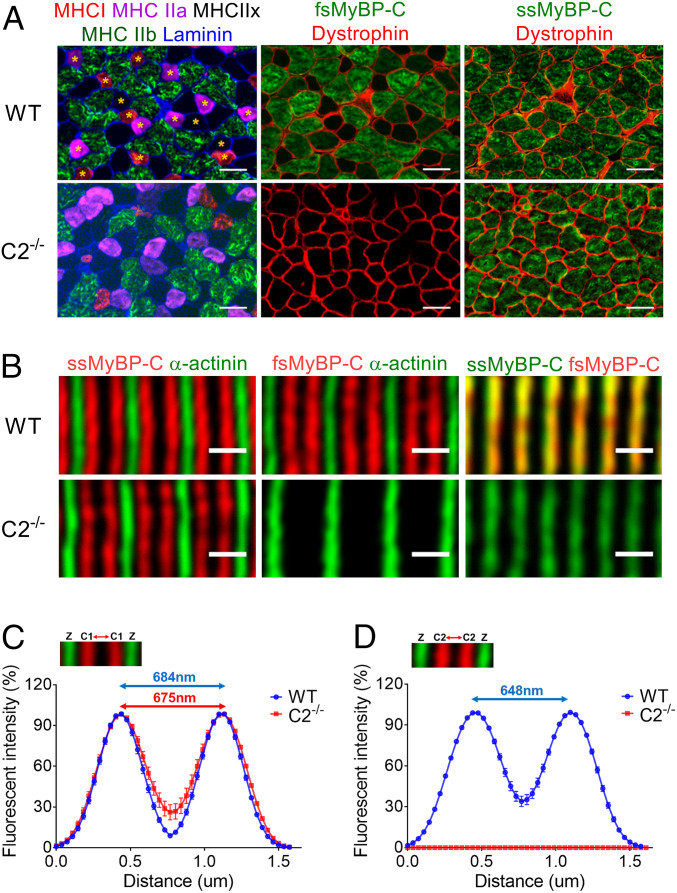
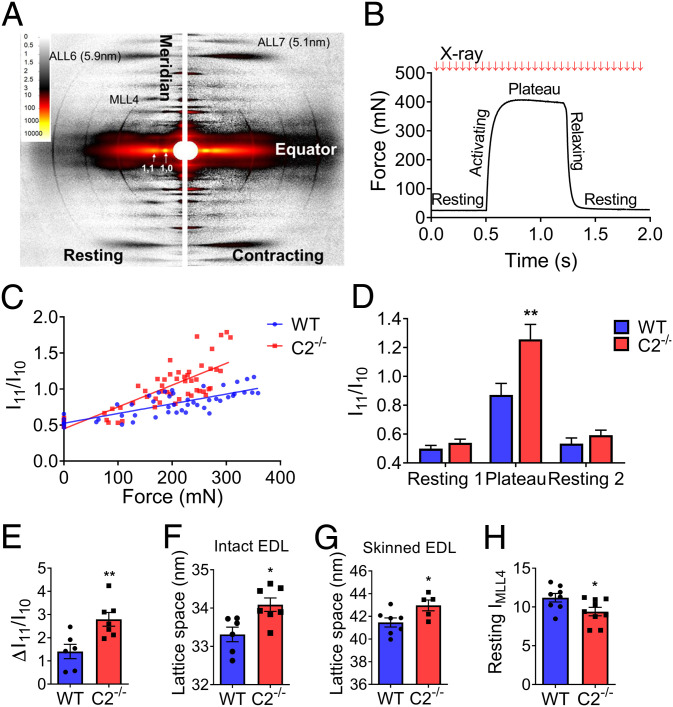
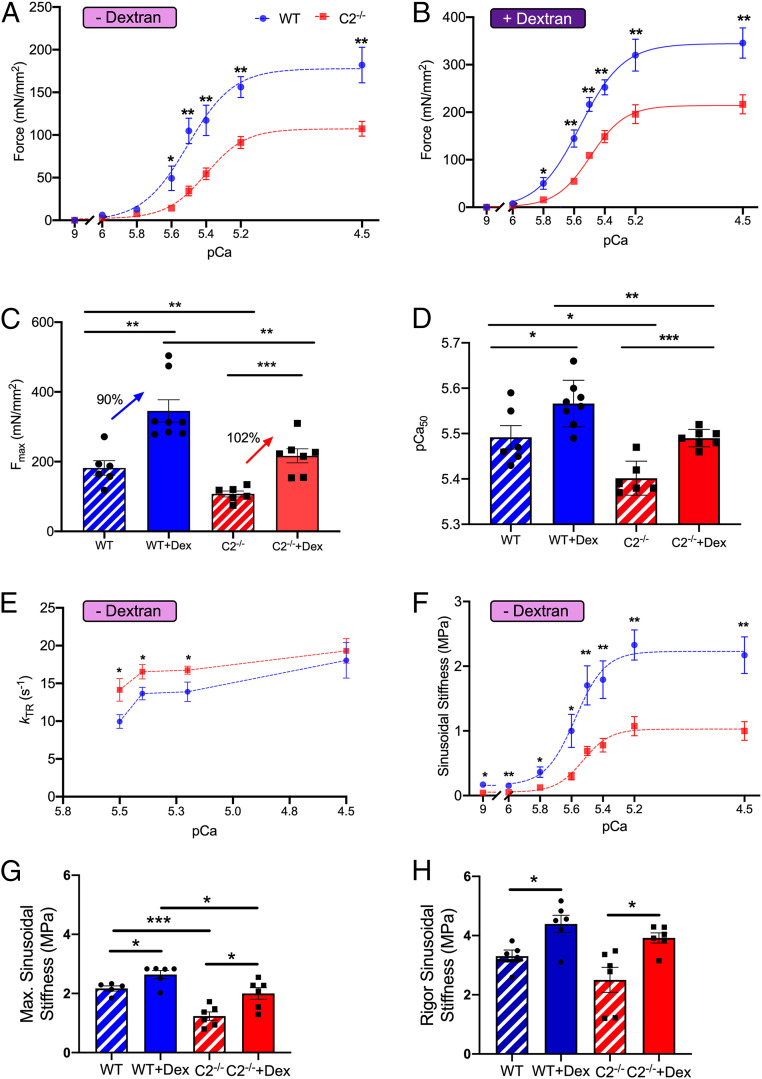
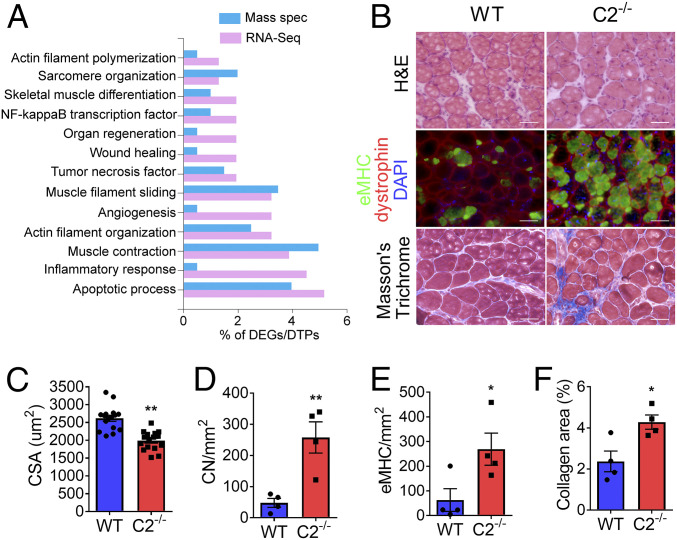
Fast skeletal myosin-binding protein-C (fMyBP-C) is one of three MyBP-C paralogs and is predominantly expressed in fast skeletal muscle. Mutations in the gene that encodes fMyBP-C, MYBPC2, are associated with distal arthrogryposis, while loss of fMyBP-C protein is associated with diseased muscle. However, the functional and structural roles of fMyBP-C in skeletal muscle remain unclear. To address this gap, we generated a homozygous fMyBP-C knockout mouse (C2-/-) and characterized it both in vivo and in vitro compared to wild-type mice. Ablation of fMyBP-C was benign in terms of muscle weight, fiber type, cross-sectional area, and sarcomere ultrastructure. However, grip strength and plantar flexor muscle strength were significantly decreased in C2-/- mice. Peak isometric tetanic force and isotonic speed of contraction were significantly reduced in isolated extensor digitorum longus (EDL) from C2-/- mice. Small-angle X-ray diffraction of C2-/- EDL muscle showed significantly increased equatorial intensity ratio during contraction, indicating a greater shift of myosin heads toward actin, while MLL4 layer line intensity was decreased at rest, indicating less ordered myosin heads. Interfilament lattice spacing increased significantly in C2-/- EDL muscle. Consistent with these findings, we observed a significant reduction of steady-state isometric force during Ca2+-activation, decreased myofilament calcium sensitivity, and sinusoidal stiffness in skinned EDL muscle fibers from C2-/- mice. Finally, C2-/- muscles displayed disruption of inflammatory and regenerative pathways, along with increased muscle damage upon mechanical overload. Together, our data suggest that fMyBP-C is essential for maximal speed and force of contraction, sarcomere integrity, and calcium sensitivity in fast-twitch muscle.
Keywords: MYBPC2; contraction; distal arthrogryposis; fMyBP-C; skeletal muscle.
Conflict of interest statement
Competing interest statement: S.S. provided consulting and collaborative research studies to the Leducq Foundation, Red Saree Inc., Greater Cincinnati Tamil Sangam, AstraZeneca, MyoKardia, Merck, and Amgen, but such work is unrelated to the content of this manuscript.
Figures
References
-
- Starr R., Offer G., Polypeptide chains of intermediate molecular weight in myosin preparations. FEBS Lett. 15, 40–44 (1971). - PubMed
-
- Gautel M., Fürst D. O., Cocco A., Schiaffino S., Isoform transitions of the myosin binding protein C family in developing human and mouse muscles: Lack of isoform transcomplementation in cardiac muscle. Circ. Res. 82, 124–129 (1998). - PubMed
-
- Flashman E., Redwood C., Moolman-Smook J., Watkins H., Cardiac myosin binding protein C: Its role in physiology and disease. Circ. Res. 94, 1279–1289 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
