

Preventing Engrailed-1 activation in fibroblasts yields wound regeneration without scarring
- PMID: 33888614
- PMCID: PMC9008875
- DOI: 10.1126/science.aba2374
Preventing Engrailed-1 activation in fibroblasts yields wound regeneration without scarring
Abstract
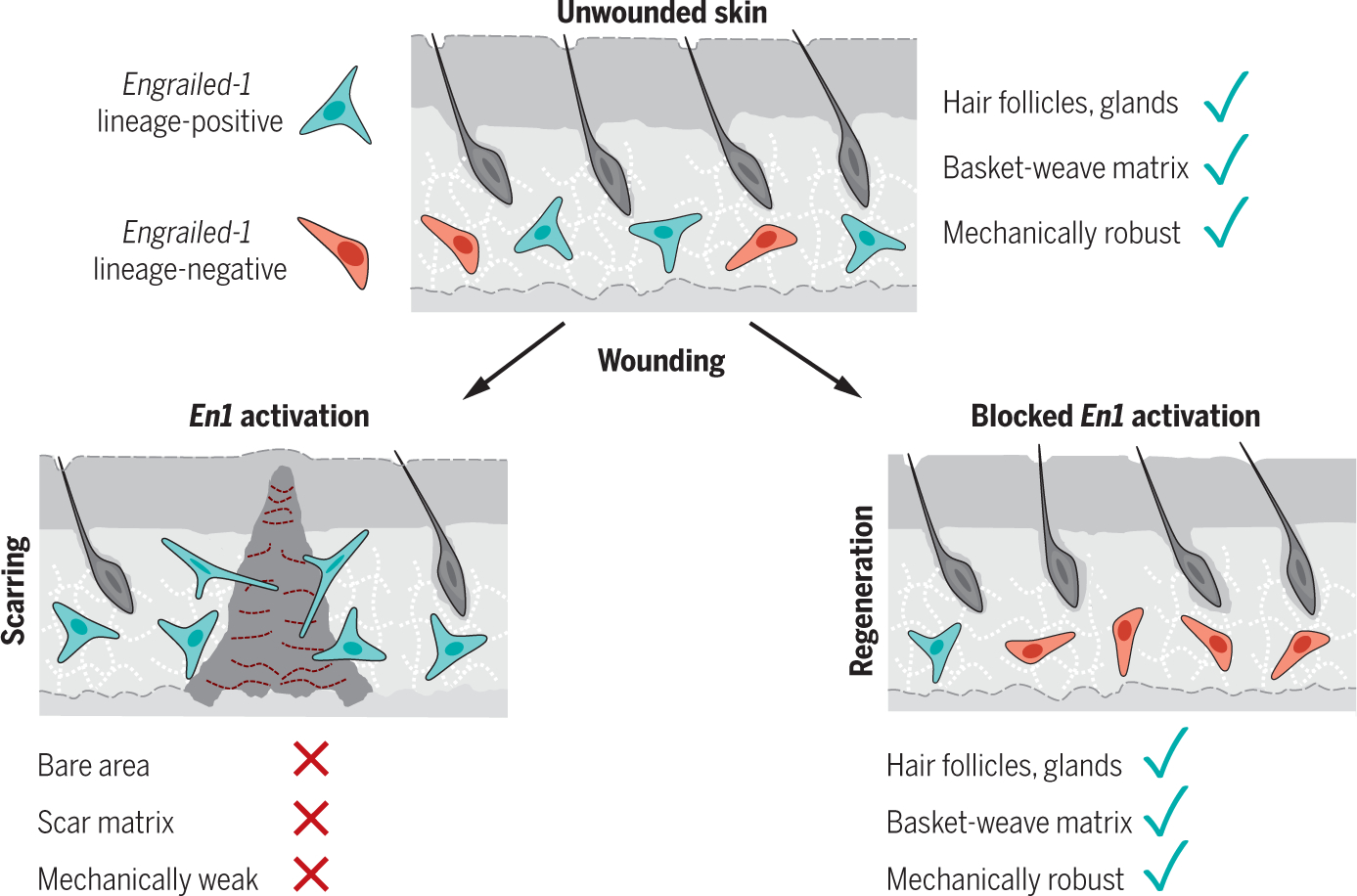
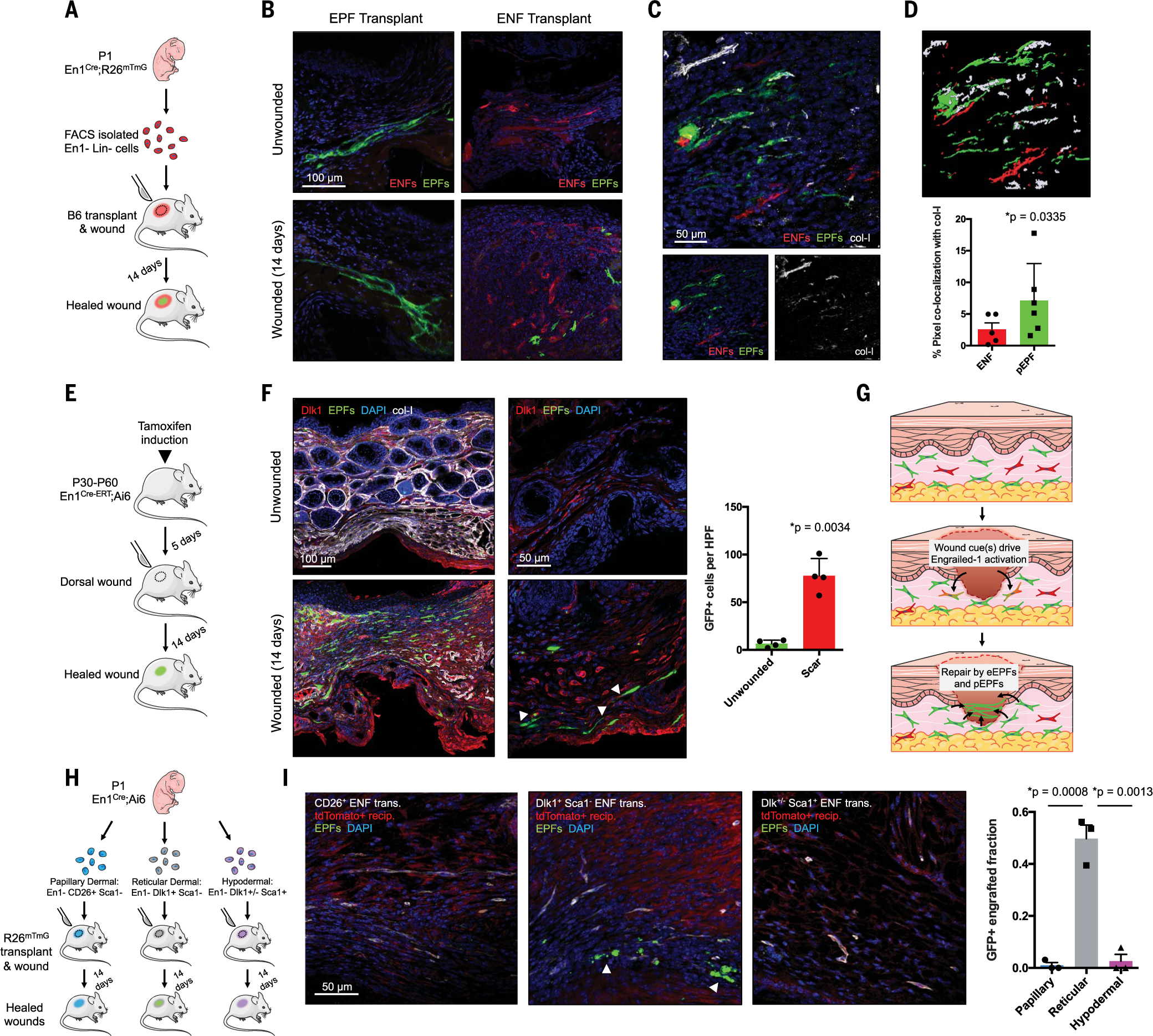
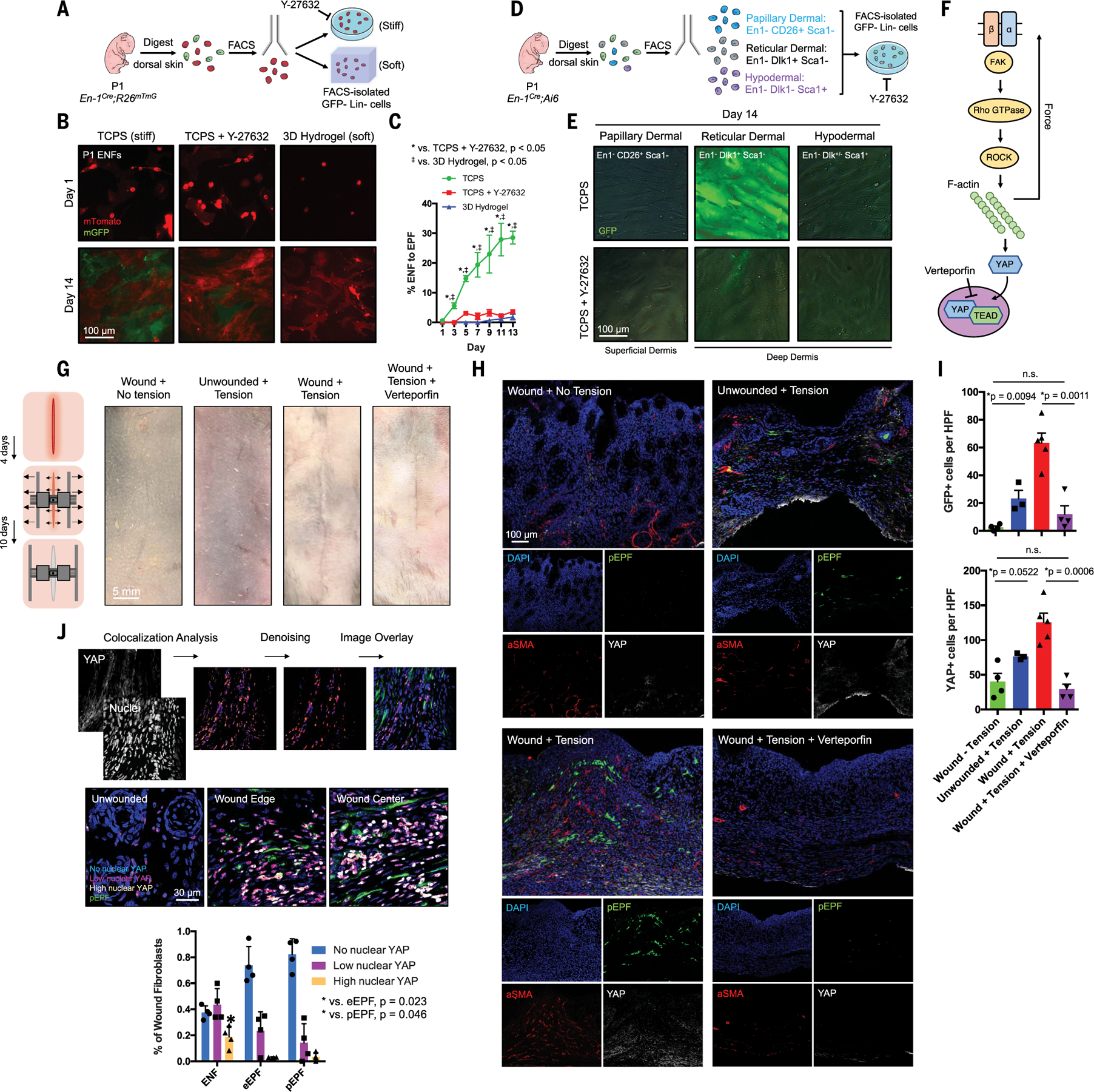
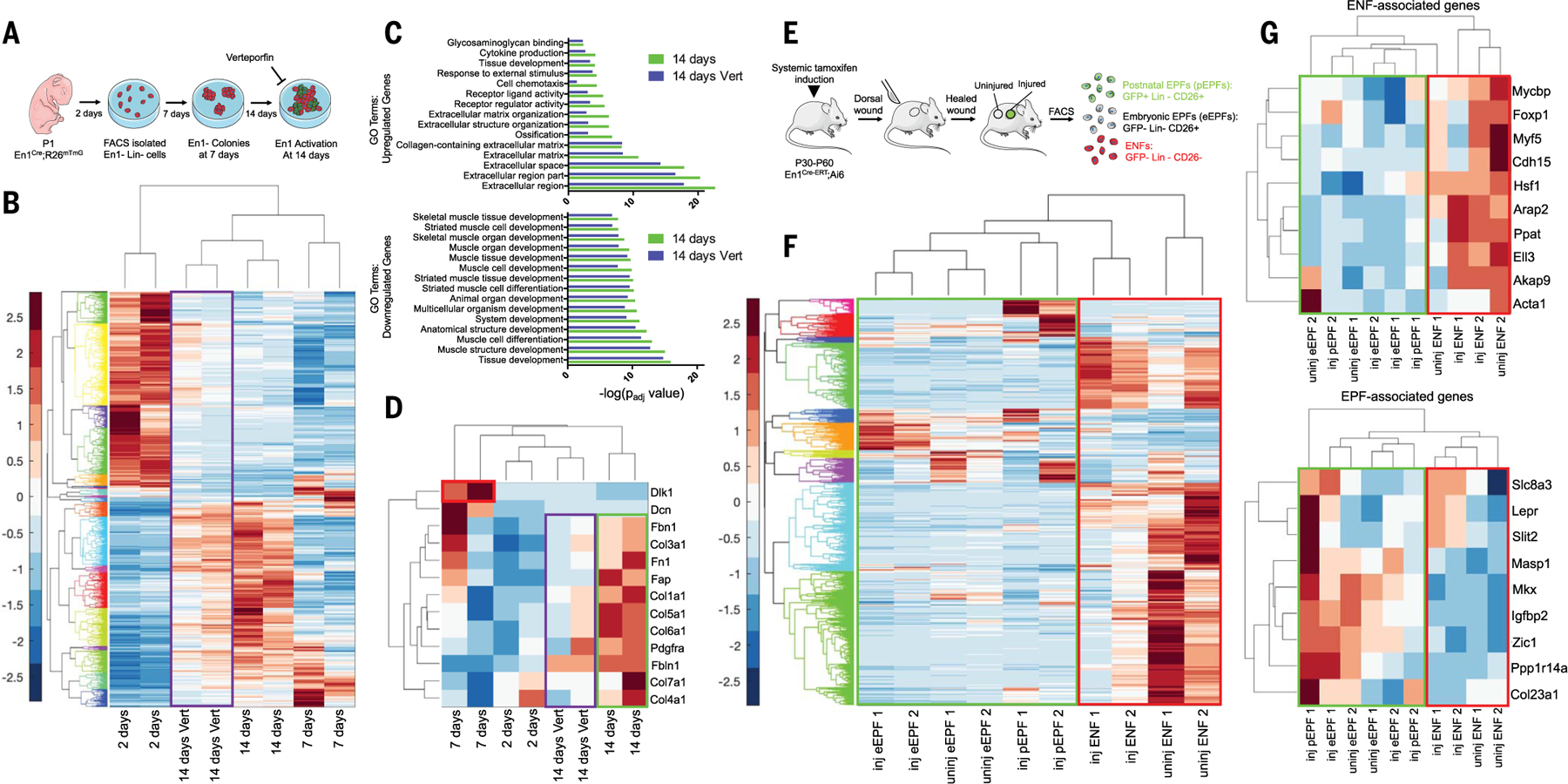
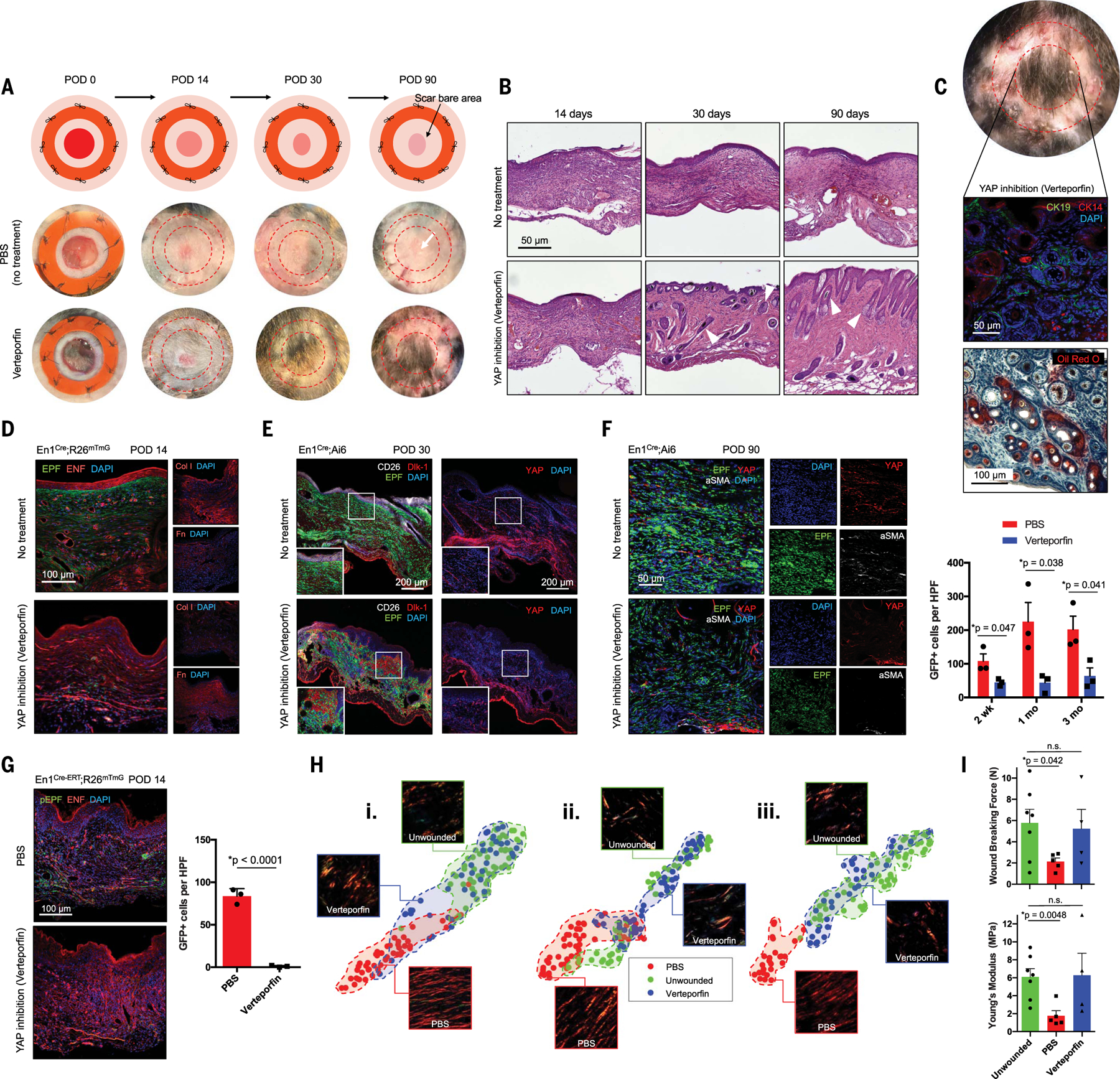
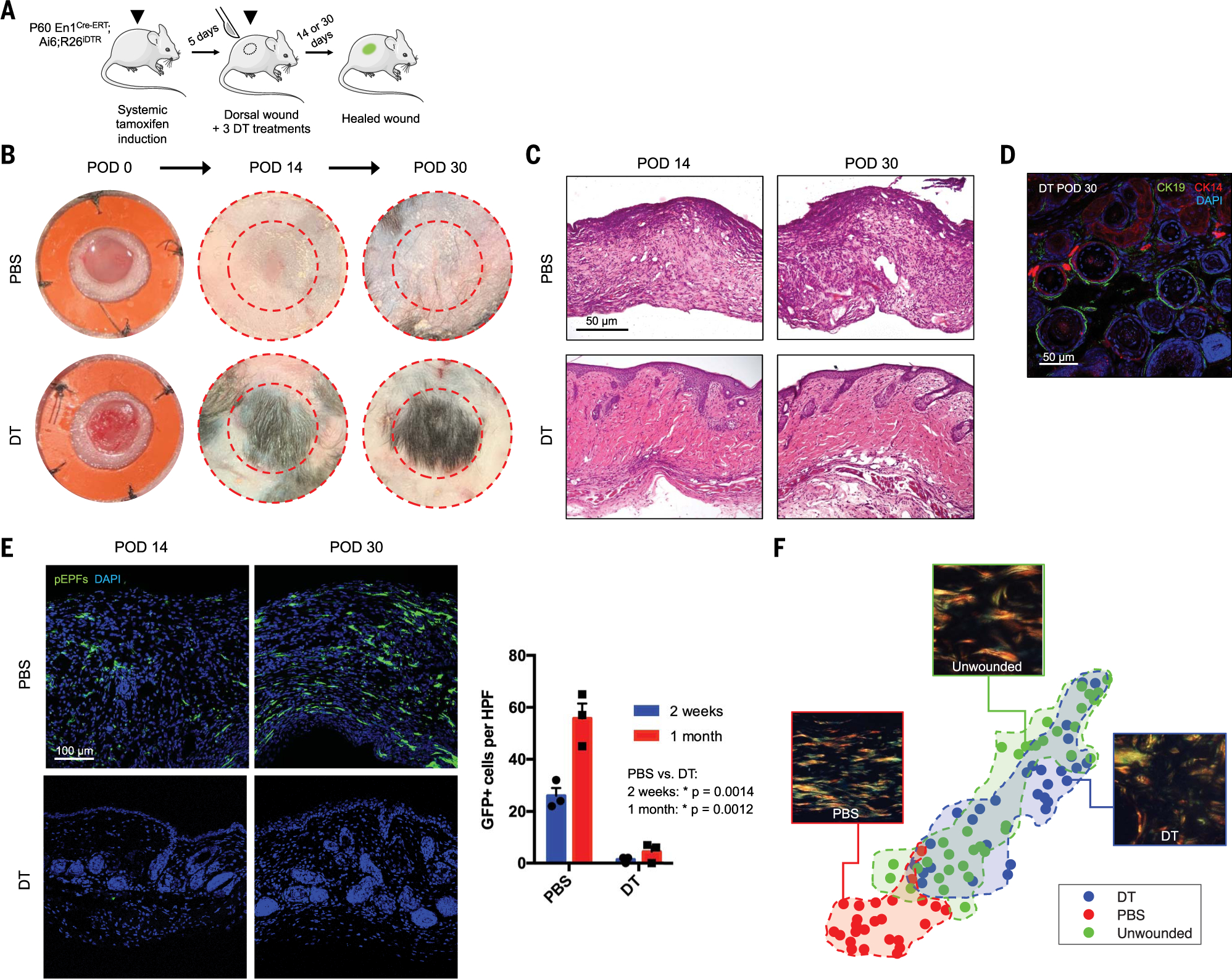
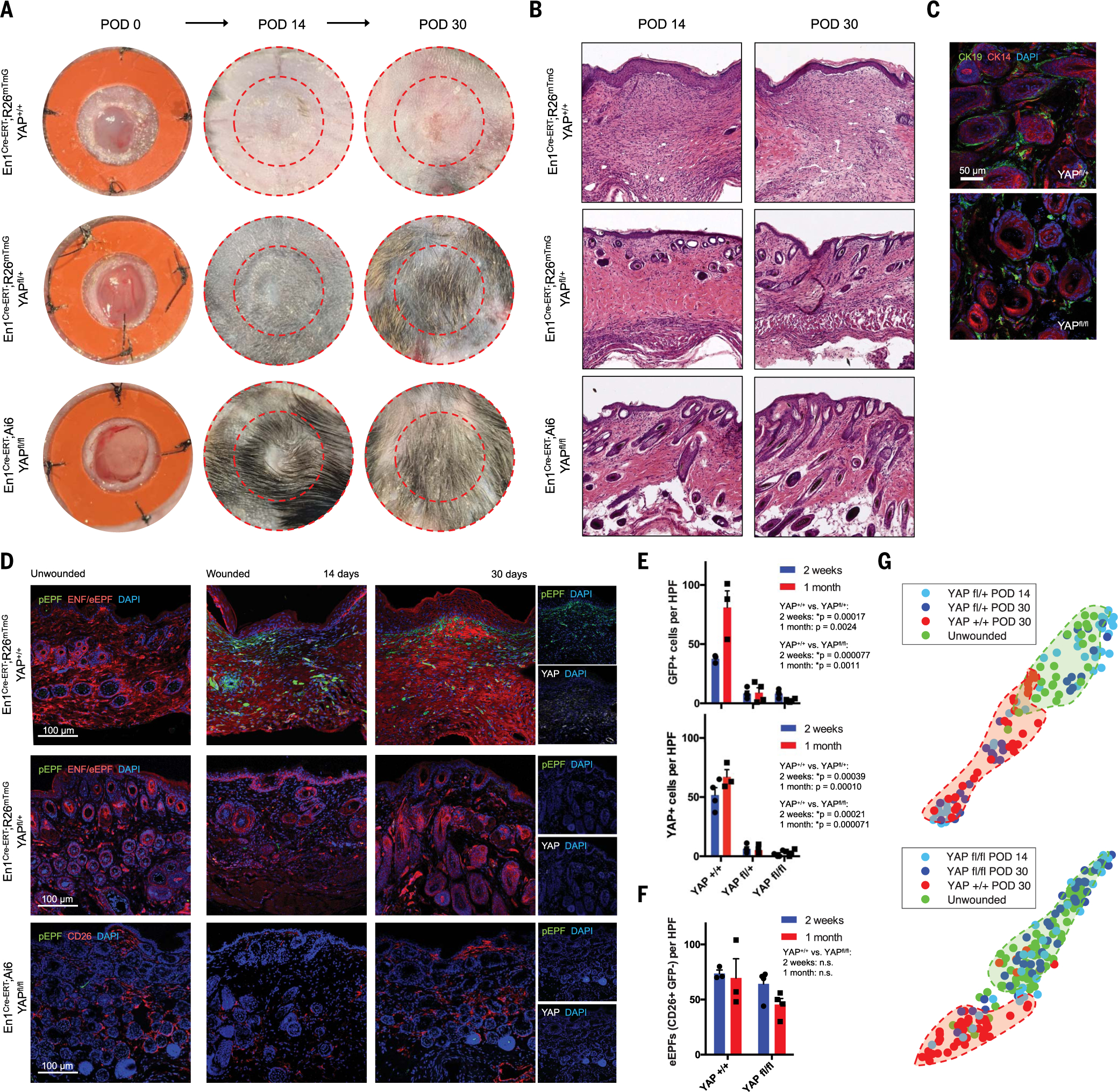
Skin scarring, the end result of adult wound healing, is detrimental to tissue form and function. Engrailed-1 lineage-positive fibroblasts (EPFs) are known to function in scarring, but Engrailed-1 lineage-negative fibroblasts (ENFs) remain poorly characterized. Using cell transplantation and transgenic mouse models, we identified a dermal ENF subpopulation that gives rise to postnatally derived EPFs by activating Engrailed-1 expression during adult wound healing. By studying ENF responses to substrate mechanics, we found that mechanical tension drives Engrailed-1 activation via canonical mechanotransduction signaling. Finally, we showed that blocking mechanotransduction signaling with either verteporfin, an inhibitor of Yes-associated protein (YAP), or fibroblast-specific transgenic YAP knockout prevents Engrailed-1 activation and promotes wound regeneration by ENFs, with recovery of skin appendages, ultrastructure, and mechanical strength. This finding suggests that there are two possible outcomes to postnatal wound healing: a fibrotic response (EPF-mediated) and a regenerative response (ENF-mediated).
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Healing without scarring.Science. 2021 Apr 23;372(6540):346-347. doi: 10.1126/science.abi5770. Science. 2021. PMID: 33888629 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
