

Replication timing maintains the global epigenetic state in human cells
- PMID: 33888635
- PMCID: PMC8173839
- DOI: 10.1126/science.aba5545
Replication timing maintains the global epigenetic state in human cells
Abstract
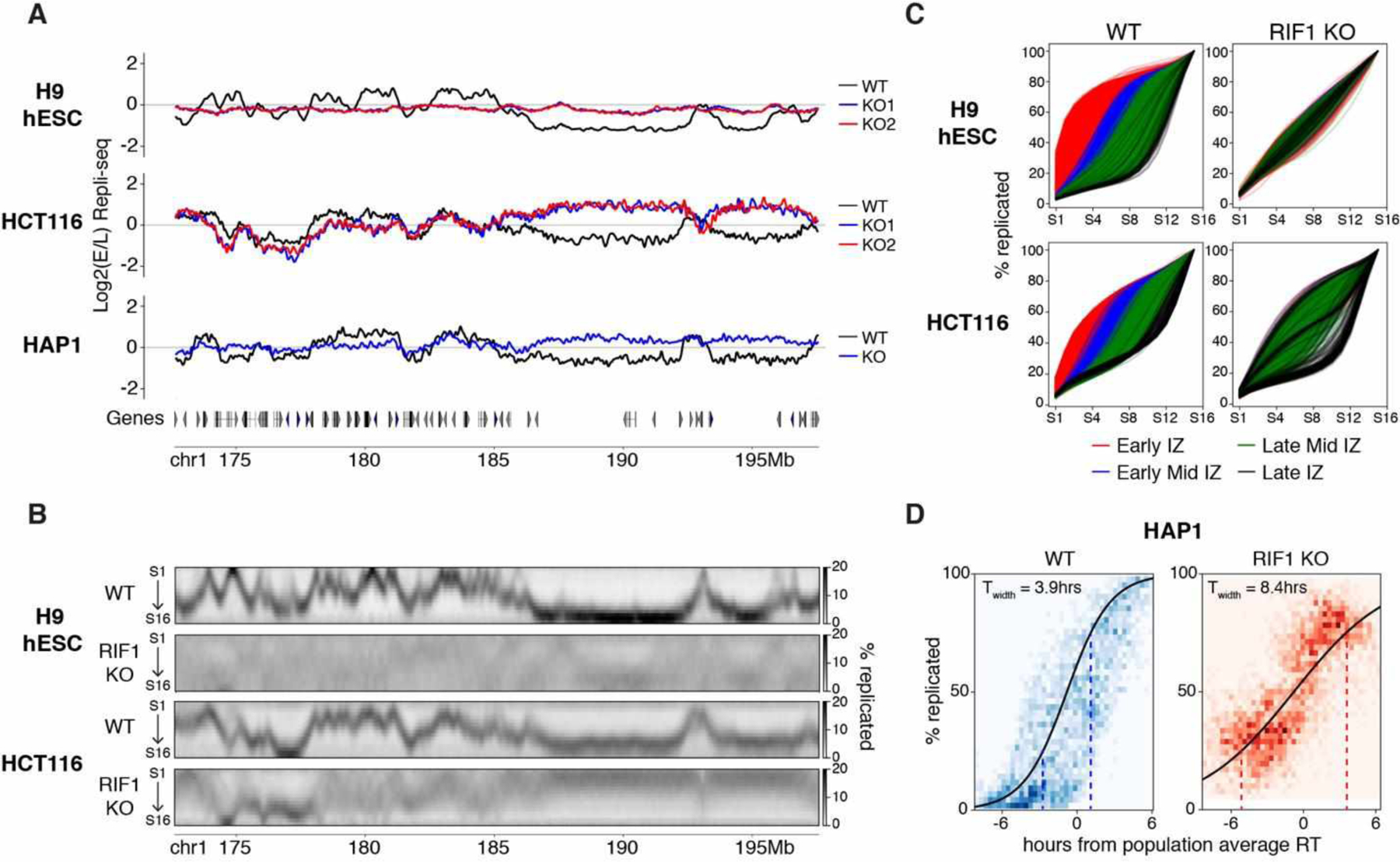
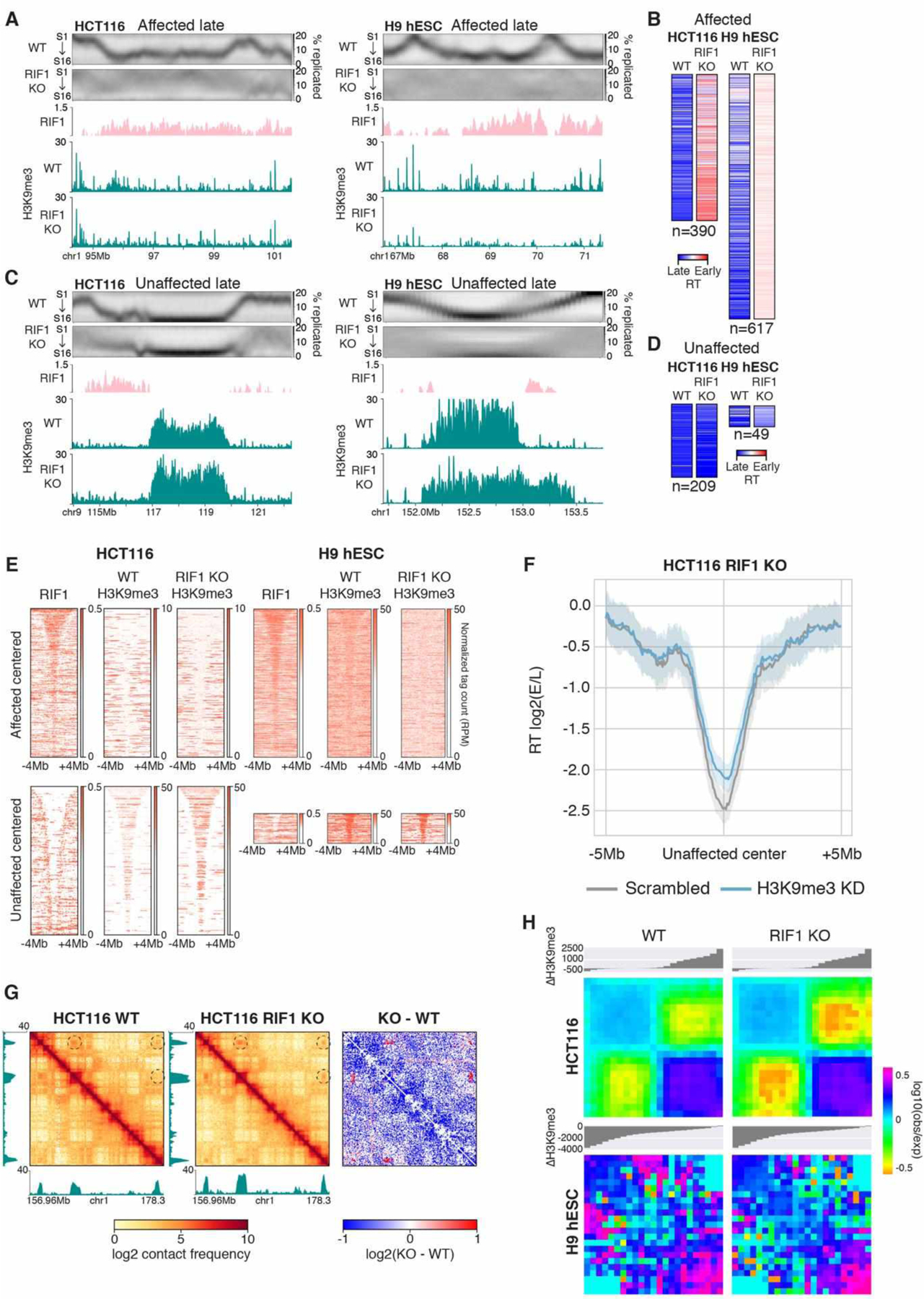
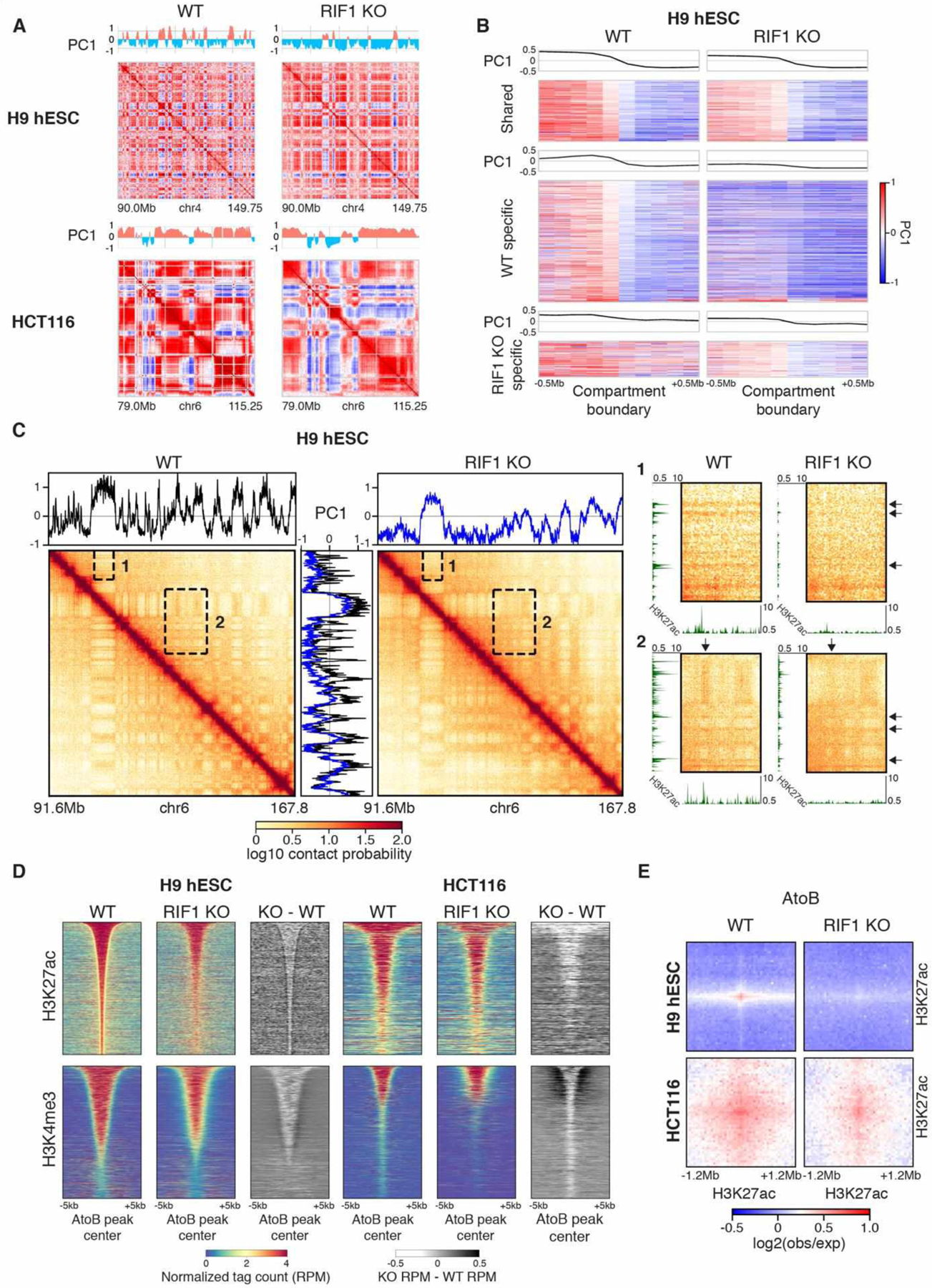
The temporal order of DNA replication [replication timing (RT)] is correlated with chromatin modifications and three-dimensional genome architecture; however, causal links have not been established, largely because of an inability to manipulate the global RT program. We show that loss of RIF1 causes near-complete elimination of the RT program by increasing heterogeneity between individual cells. RT changes are coupled with widespread alterations in chromatin modifications and genome compartmentalization. Conditional depletion of RIF1 causes replication-dependent disruption of histone modifications and alterations in genome architecture. These effects were magnified with successive cycles of altered RT. These results support models in which the timing of chromatin replication and thus assembly plays a key role in maintaining the global epigenetic state.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures


References
-
- Reverón-Gómez N, González-Aguilera C, Stewart-Morgan KR, Petryk N, Flury V, Graziano S, Johansen JV, Jakobsen JS, Alabert C, Groth A, Accurate Recycling of Parental Histones Reproduces the Histone Modification Landscape during DNA Replication. Mol. Cell (2018), doi:10.1016/j.molcel.2018.08.010. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
