

Environmental connectivity controls diversity in soil microbial communities
- PMID: 33888858
- PMCID: PMC8062517
- DOI: 10.1038/s42003-021-02023-2
Environmental connectivity controls diversity in soil microbial communities
Abstract
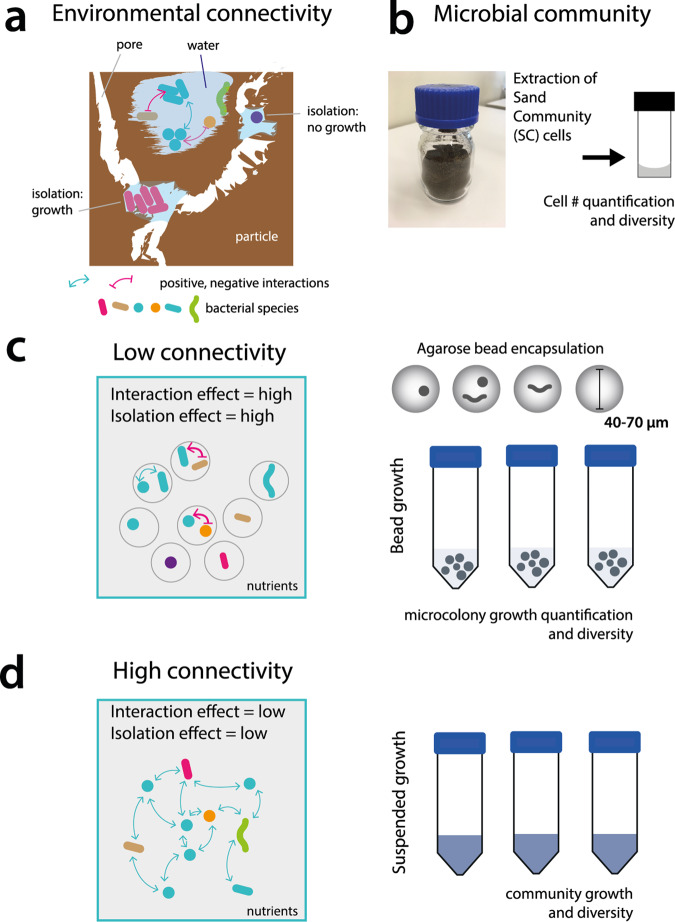
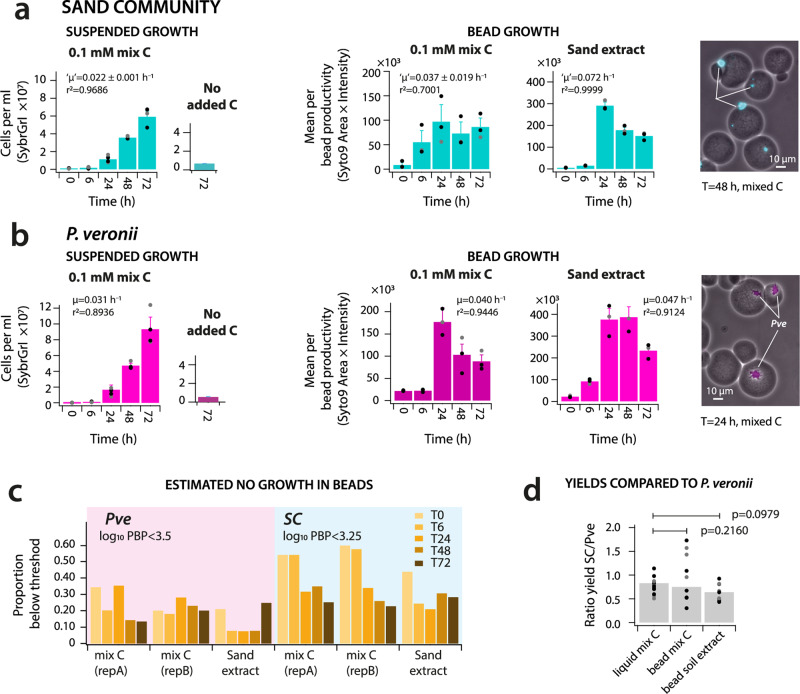
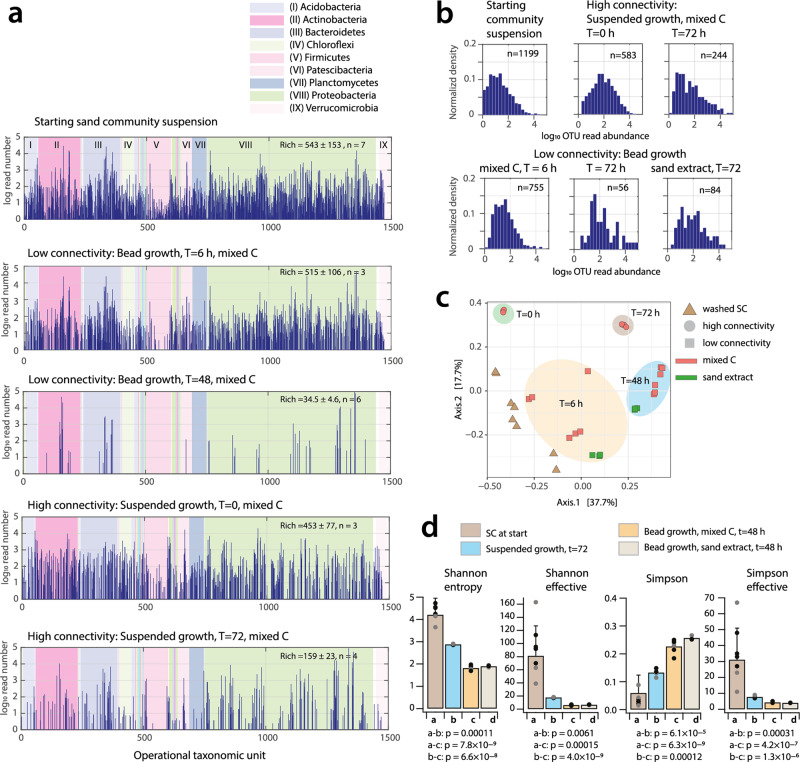
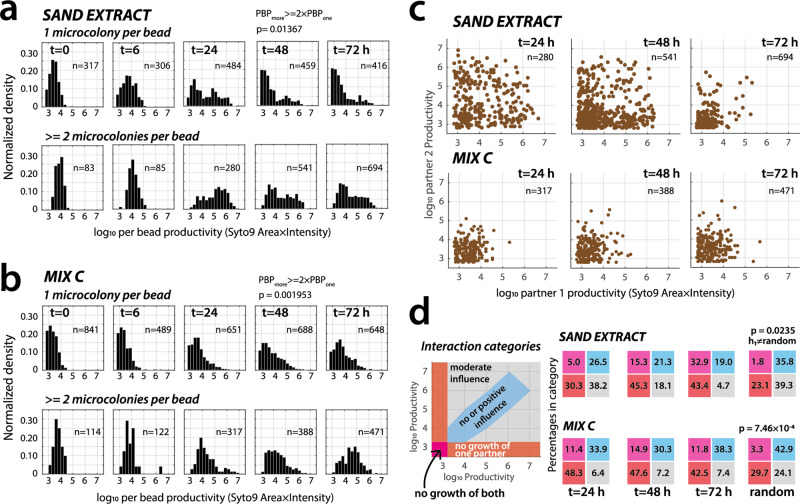
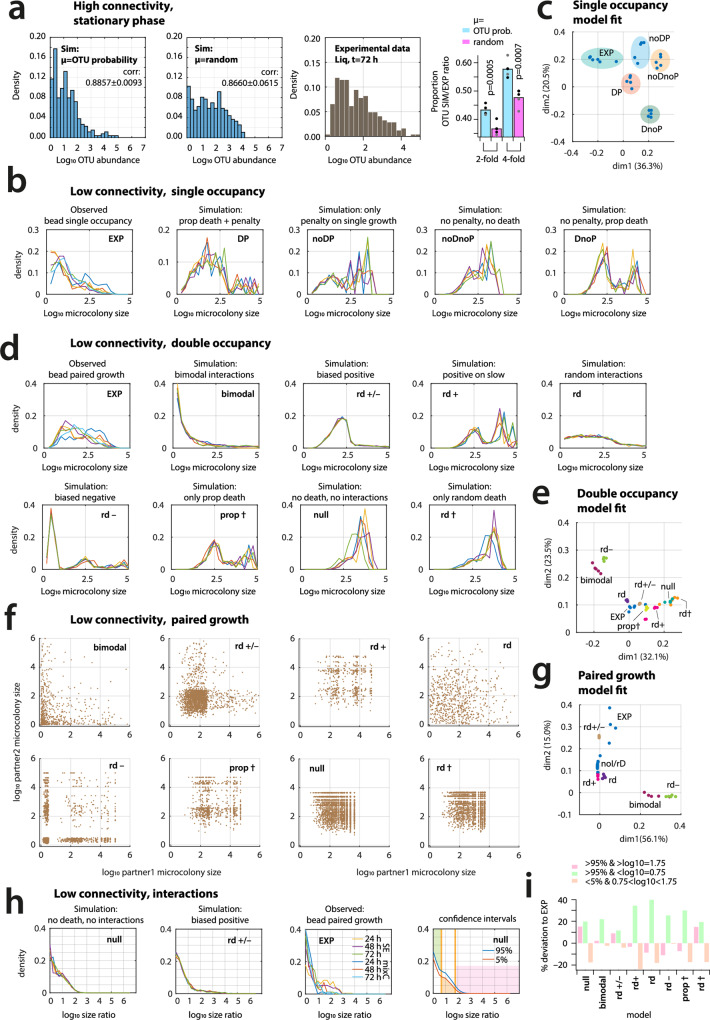
Interspecific interactions are thought to govern the stability and functioning of microbial communities, but the influence of the spatial environment and its structural connectivity on the potential of such interactions to unfold remain largely unknown. Here we studied the effects on community growth and microbial diversity as a function of environmental connectivity, where we define environmental connectivity as the degree of habitat fragmentation preventing microbial cells from living together. We quantitatively compared growth of a naturally-derived high microbial diversity community from soil in a completely mixed liquid suspension (high connectivity) to growth in a massively fragmented and poorly connected environment (low connectivity). The low connectivity environment consisted of homogenously-sized miniature agarose beads containing random single or paired founder cells. We found that overall community growth was the same in both environments, but the low connectivity environment dramatically reduced global community-level diversity compared to the high connectivity environment. Experimental observations were supported by community growth modeling. The model predicts a loss of diversity in the low connectivity environment as a result of negative interspecific interactions becoming more dominant at small founder species numbers. Counterintuitively for the low connectivity environment, growth of isolated single genotypes was less productive than that of random founder genotype cell pairs, suggesting that the community as a whole profited from emerging positive interspecific interactions. Our work demonstrates the importance of environmental connectivity for growth of natural soil microbial communities, which aids future efforts to intervene in or restore community composition to achieve engineering and biotechnological objectives.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
