

Serotonergic Modulation of Persistent Inward Currents in Serotonergic Neurons of Medulla in ePet-EYFP Mice
- PMID: 33889077
- PMCID: PMC8055846
- DOI: 10.3389/fncir.2021.657445
Serotonergic Modulation of Persistent Inward Currents in Serotonergic Neurons of Medulla in ePet-EYFP Mice
Abstract
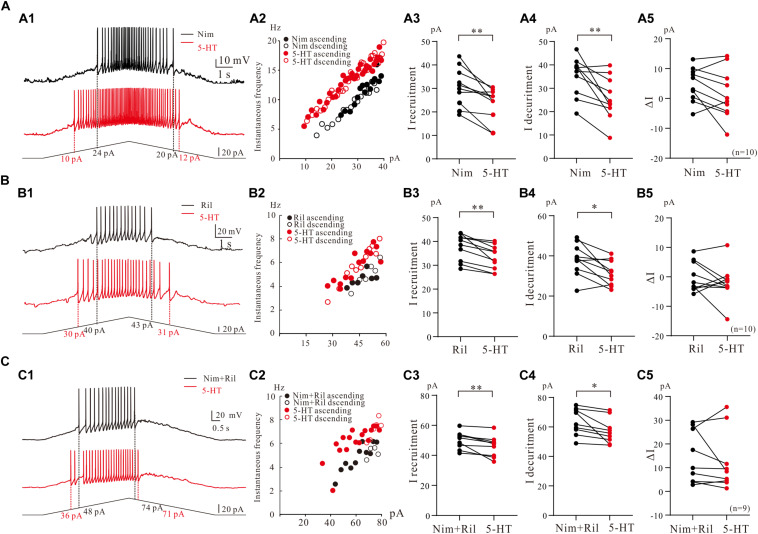
Serotonergic (5-HT) neurons in the medulla play multiple functional roles associated with many symptoms and motor activities. The descending serotonergic pathway from medulla is essential for initiating locomotion. However, the ionic properties of 5-HT neurons in the medulla remain unclear. Using whole-cell patch-clamp technique, we studied the biophysical and modulatory properties of persistent inward currents (PICs) in 5-HT neurons of medulla in ePet-EYFP transgenic mice (P3-P6). PICs were recorded by a family of voltage bi-ramps (10-s duration, 40-mV peak step), and the ascending and descending PICs were mirrored to analyze the PIC hysteresis. PICs were found in 77% of 5-HT neurons (198/258) with no significant difference between parapyramidal region (n = 107) and midline raphe nuclei (MRN) (n = 91) in either PIC onset (-47.4 ± 10 mV and -48.7 ± 7 mV; P = 0.44) or PIC amplitude (226.9 ± 138 pA and 259.2 ± 141 pA; P = 0.29). Ninety-six percentage (191/198) of the 5-HT neurons displayed counterclockwise hysteresis and four percentage (7/198) exhibited the clockwise hysteresis. The composite PICs could be differentiated as calcium component (Ca_PIC) by bath application of nimodipine (25 μM), sodium component (Na_PIC) by tetrodotoxin (TTX, 2 μM), and TTX- and dihydropyridine-resistance component (TDR_PIC) by TTX and nimodipine. Ca_PIC, Na_PIC and TDR_PIC all contributed to upregulation of excitability of 5-HT neurons. 5-HT (15 μM) enhanced the PICs, including a 26% increase in amplitude of the compound currents of Ca_PIC and TDR_PIC (P < 0.001, n = 9), 3.6 ± 5 mV hyperpolarization of Na_PIC and TDR_PIC onset (P < 0.05, n = 12), 30% increase in amplitude of TDR_PIC (P < 0.01), and 2.0 ± 3 mV hyperpolarization of TDR_PIC onset (P < 0.05, n = 18). 5-HT also facilitated repetitive firing of 5-HT neurons through modulation of composite PIC, Na_PIC and TDR_PIC, and Ca_PIC and TDR_PIC, respectively. In particular, the high voltage-activated TDR_PIC facilitated the repetitive firing in higher membrane potential, and this facilitation could be amplified by 5-HT. Morphological data analysis indicated that the dendrites of 5-HT neurons possessed dense spherical varicosities intensively crossing 5-HT neurons in medulla. We characterized the PICs in 5-HT neurons and unveiled the mechanism underlying upregulation of excitability of 5-HT neurons through serotonergic modulation of PICs. This study provided insight into channel mechanisms responsible for the serotonergic modulation of serotonergic neurons in brainstem.
Keywords: medulla; neuromodulation; neuronal excitability; persistent inward currents; serotonergic neurons.
Copyright © 2021 Cheng, Song, Ge and Dai.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials