

Human placental cytotrophoblast epigenome dynamics over gestation and alterations in placental disease
- PMID: 33891899
- PMCID: PMC8650129
- DOI: 10.1016/j.devcel.2021.04.001
Human placental cytotrophoblast epigenome dynamics over gestation and alterations in placental disease
Abstract
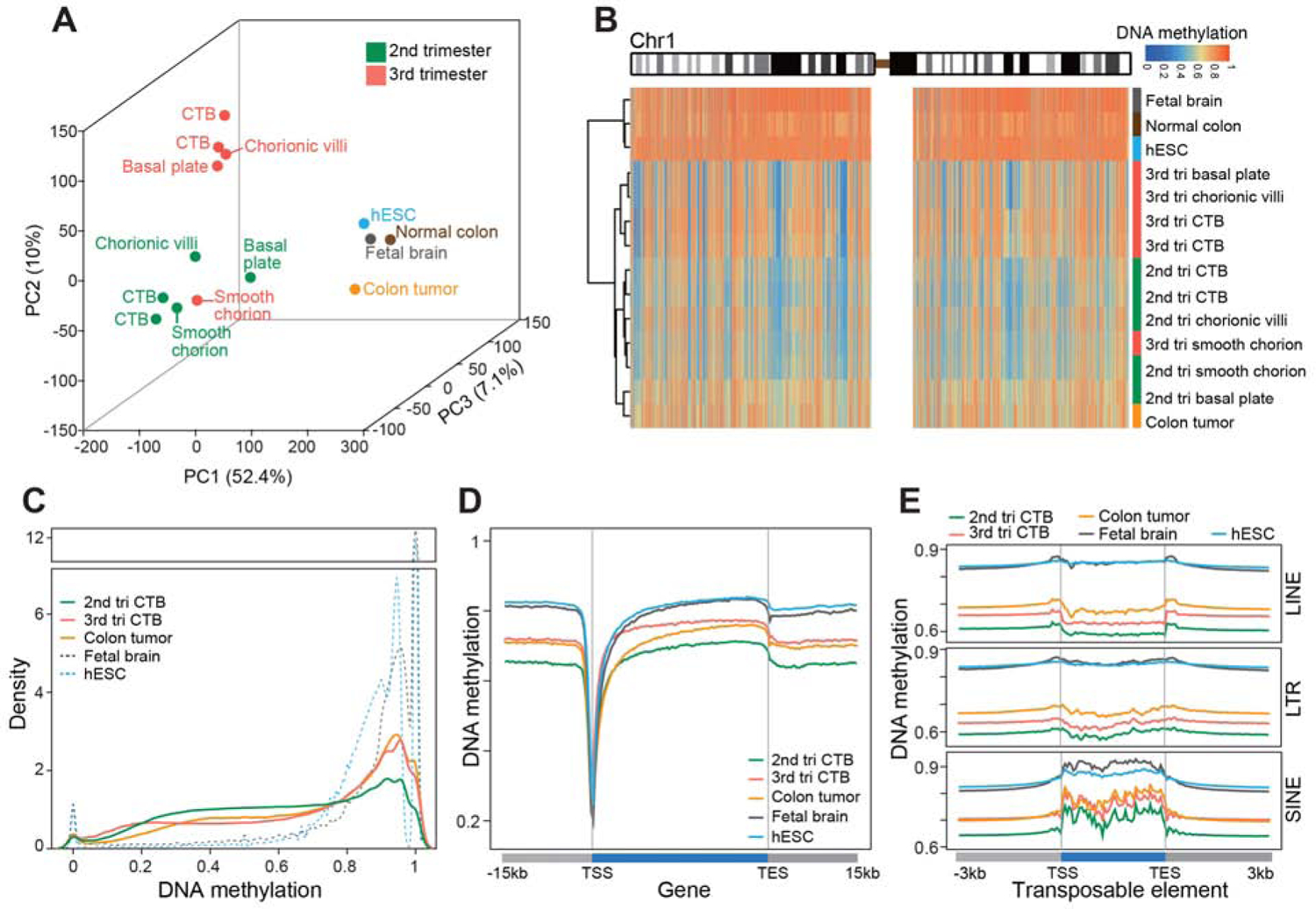
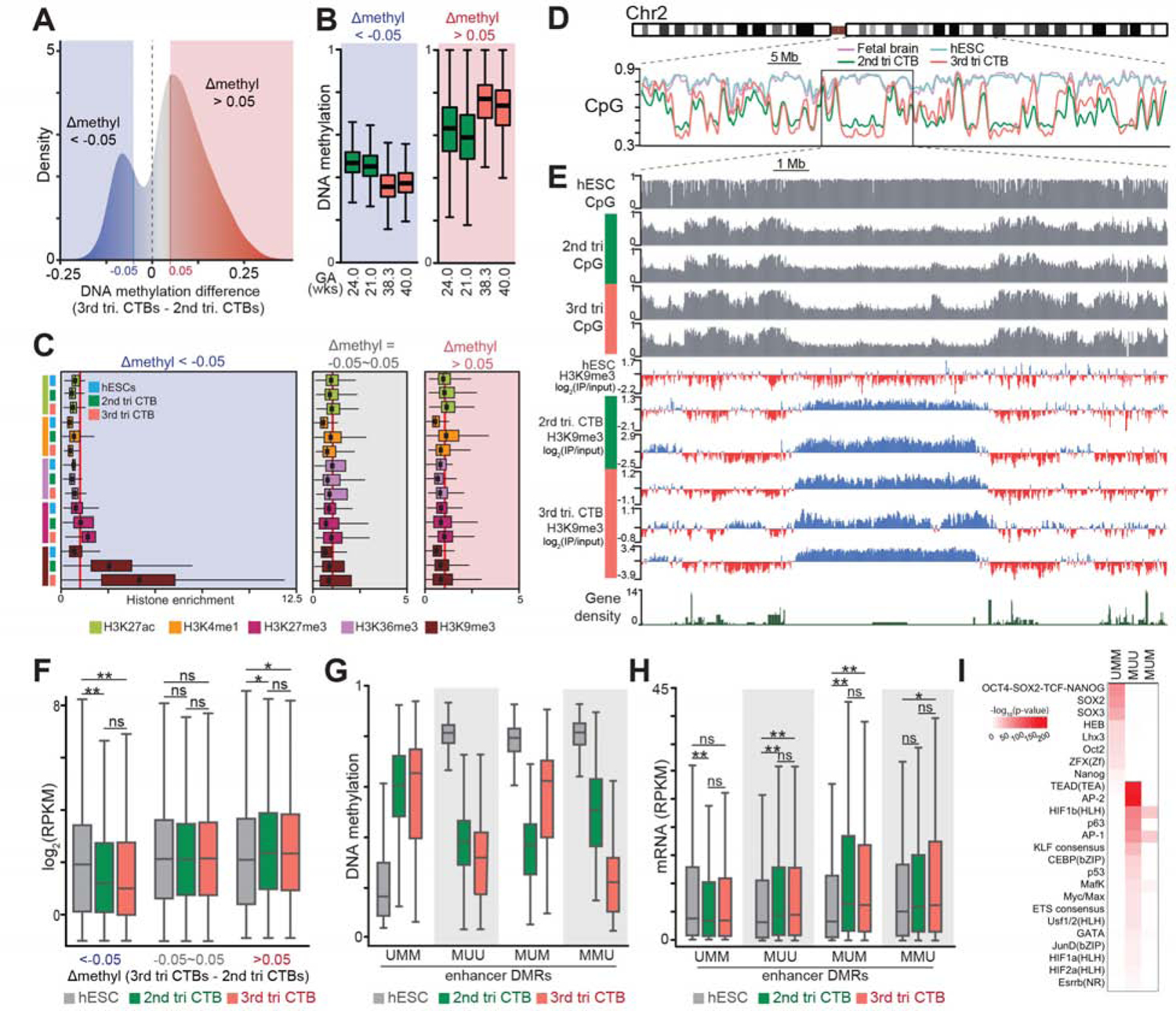
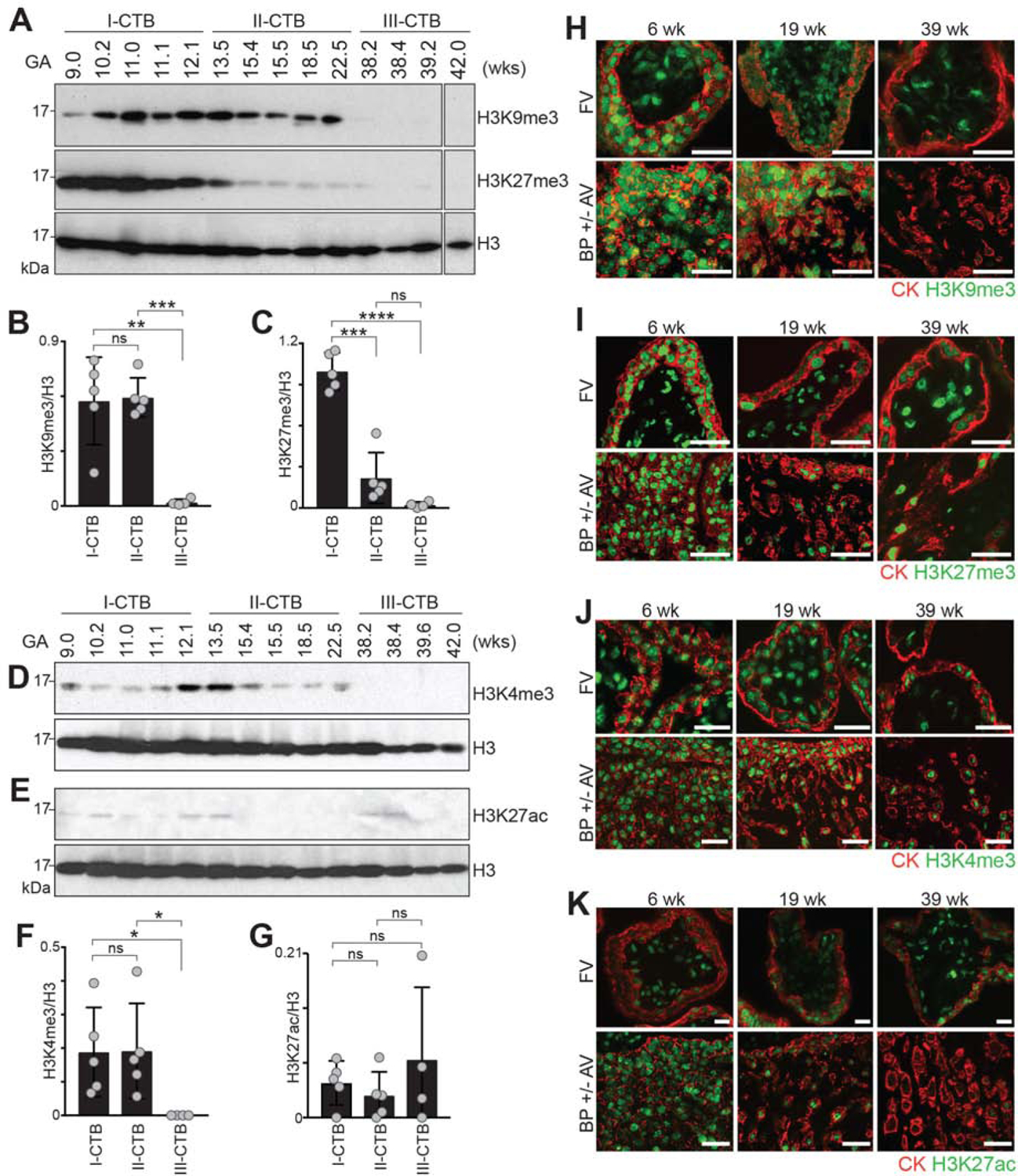
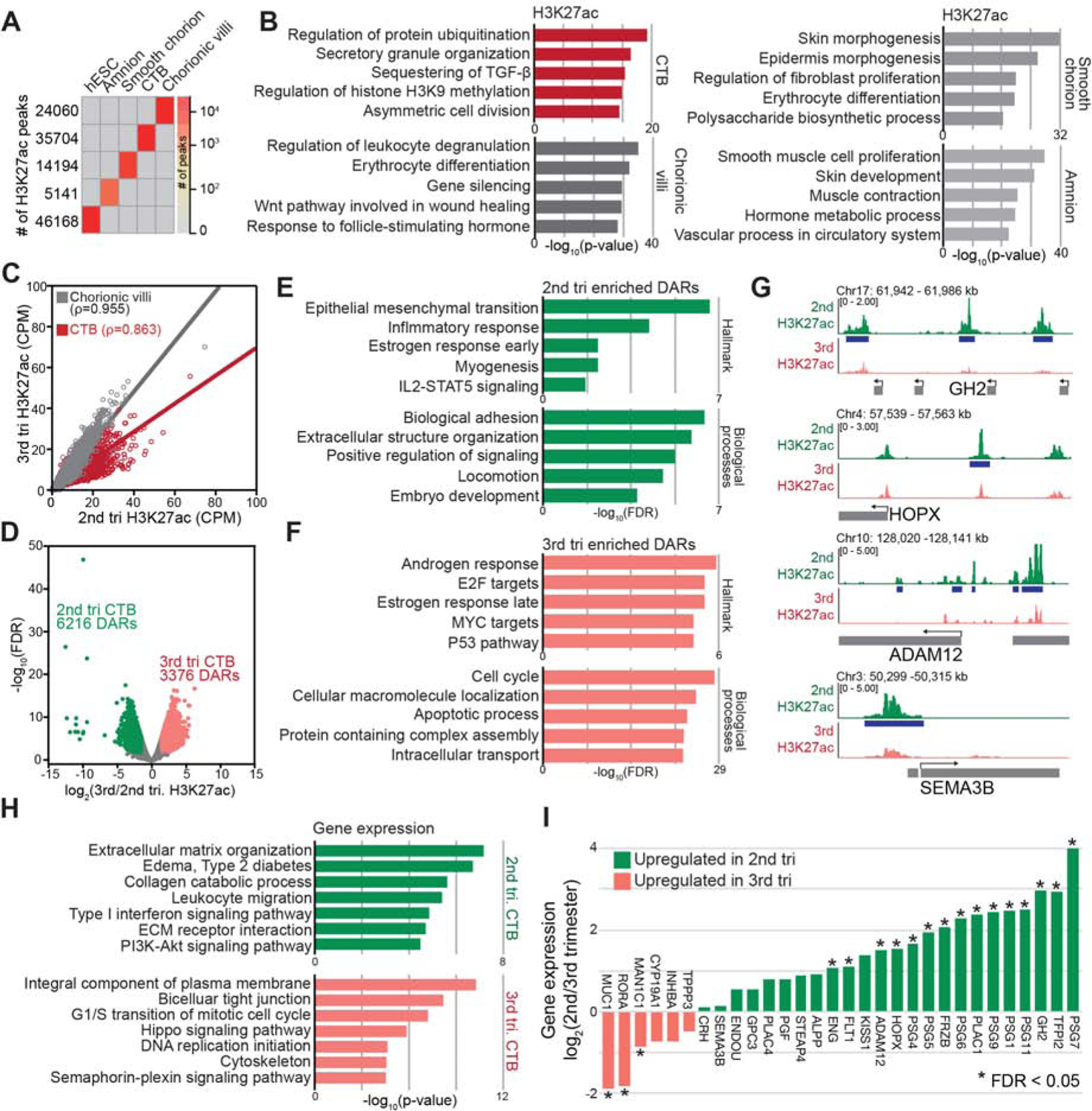
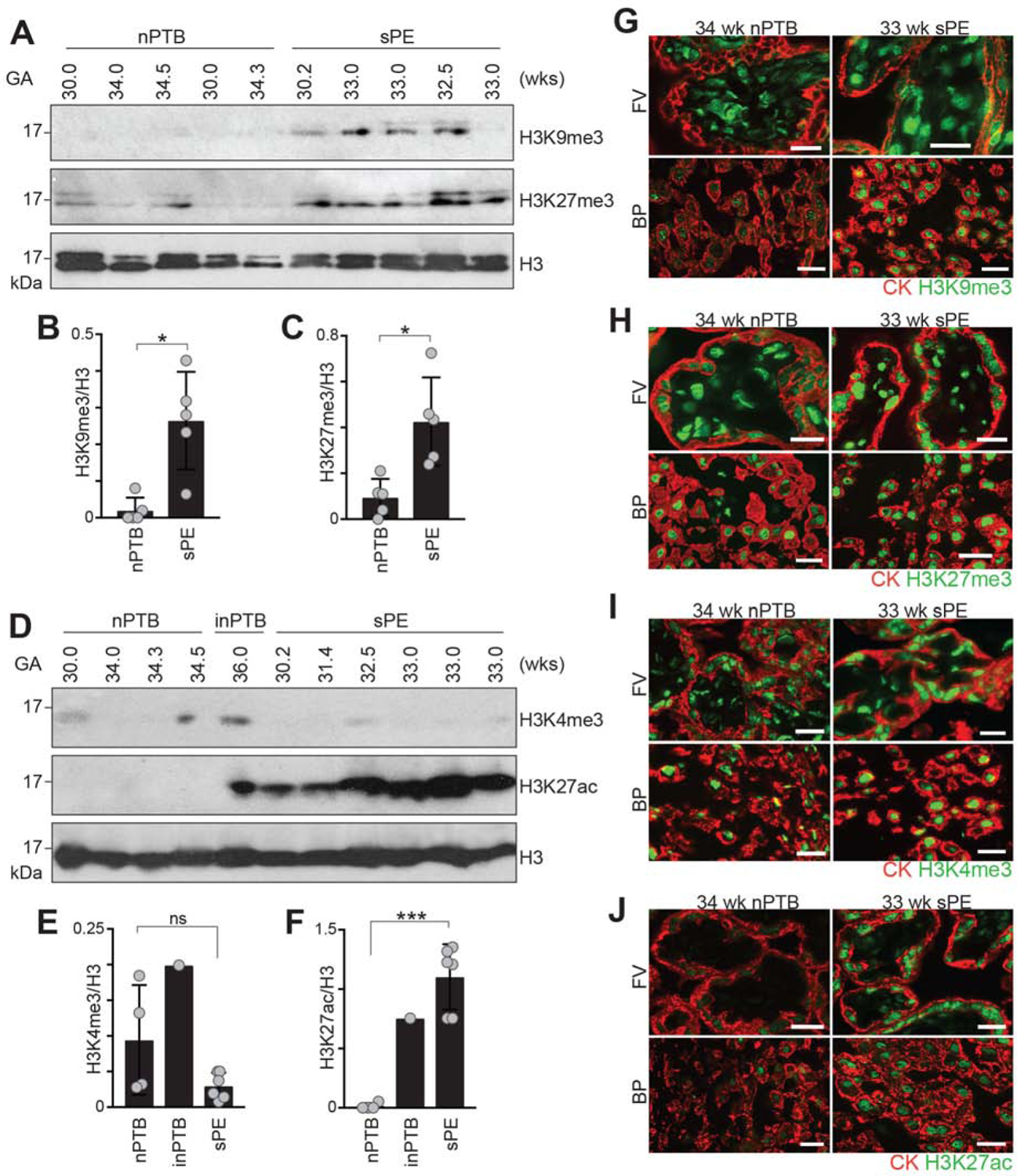
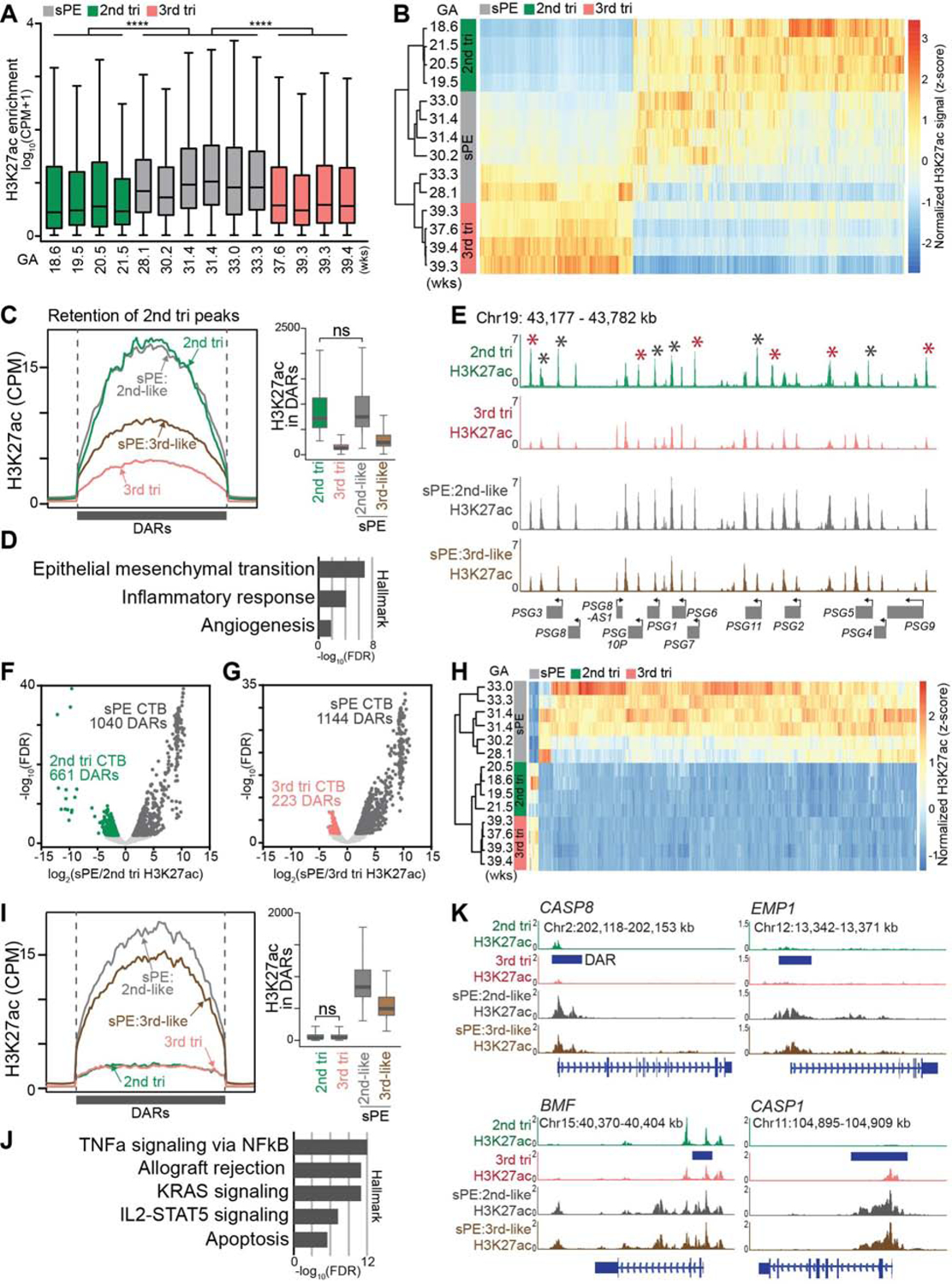
The human placenta and its specialized cytotrophoblasts rapidly develop, have a compressed lifespan, govern pregnancy outcomes, and program the offspring's health. Understanding the molecular underpinnings of these behaviors informs development and disease. Profiling the extraembryonic epigenome and transcriptome during the 2nd and 3rd trimesters revealed H3K9 trimethylation overlapping deeply DNA hypomethylated domains with reduced gene expression and compartment-specific patterns that illuminated their functions. Cytotrophoblast DNA methylation increased, and several key histone modifications decreased across the genome as pregnancy advanced. Cytotrophoblasts from severe preeclampsia had substantially increased H3K27 acetylation globally and at genes that are normally downregulated at term but upregulated in this syndrome. In addition, some cases had an immature pattern of H3K27ac peaks, and others showed evidence of accelerated aging, suggesting subtype-specific alterations in severe preeclampsia. Thus, the cytotrophoblast epigenome dramatically reprograms during pregnancy, placental disease is associated with failures in this process, and H3K27 hyperacetylation is a feature of severe preeclampsia.
Keywords: DNA methylation; H3K27ac; H3K9me3; chorionic villi; cytotrophoblast; gestational regulation; histone modification; human placenta; placental disease; preeclampsia.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.J.F. and M.M. are consultants for Novo Nordisk. The other authors declare no competing interests.
Figures
Comment in
-
Epigenetic states in the human placenta: A singular epigenome for an exceptional tissue.Dev Cell. 2021 May 3;56(9):1211-1212. doi: 10.1016/j.devcel.2021.04.011. Dev Cell. 2021. PMID: 33945779
References
-
- ACOG Committee on Practice Bulletins--Obstetrics (2002). ACOG practice bulletin. Diagnosis and management of preeclampsia and eclampsia. Number 33, January 2002. Obstet. Gynecol 99, 159–167. - PubMed
-
- American College of Obstetricians and Gynecologists, Task Force on Hypertension in Pregnancy (2013). Hypertension in pregnancy. Report of the American College of Obstetricians and Gynecologists’ Task Force on Hypertension in Pregnancy. pp. 1122–1131. - PubMed
-
- Au Yeung WK, Brind’Amour J, Hatano Y, Yamagata K, Feil R, Lorincz MC, Tachibana M, Shinkai Y, and Sasaki H (2019). Histone H3K9 Methyltransferase G9a in Oocytes Is Essential for Preimplantation Development but Dispensable for CG Methylation Protection. Cell Rep 27, 282–293.e284. - PubMed
-
- Barker DJ, Winter PD, Osmond C, Margetts B, and Simmonds SJ (1989). Weight in infancy and death from ischaemic heart disease. Lancet 2, 577–580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U01 HG009391/HG/NHGRI NIH HHS/United States
- U01 ES017154/ES/NIEHS NIH HHS/United States
- U41 HG010972/HG/NHGRI NIH HHS/United States
- R37 HD076253/HD/NICHD NIH HHS/United States
- U01 CA200060/CA/NCI NIH HHS/United States
- R01 ES024992/ES/NIEHS NIH HHS/United States
- R25 DA027995/DA/NIDA NIH HHS/United States
- R01 HG007354/HG/NHGRI NIH HHS/United States
- R01 HD092419/HD/NICHD NIH HHS/United States
- U24 HG012070/HG/NHGRI NIH HHS/United States
- R01 HG007175/HG/NHGRI NIH HHS/United States
- P50 HD055764/HD/NICHD NIH HHS/United States
- U24 ES026699/ES/NIEHS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
