

G-quadruplexes are transcription factor binding hubs in human chromatin
- PMID: 33892767
- PMCID: PMC8063395
- DOI: 10.1186/s13059-021-02324-z
G-quadruplexes are transcription factor binding hubs in human chromatin
Abstract
Background: The binding of transcription factors (TF) to genomic targets is critical in the regulation of gene expression. Short, double-stranded DNA sequence motifs are routinely implicated in TF recruitment, but many questions remain on how binding site specificity is governed.
Results: Herein, we reveal a previously unappreciated role for DNA secondary structures as key features for TF recruitment. In a systematic, genome-wide study, we discover that endogenous G-quadruplex secondary structures (G4s) are prevalent TF binding sites in human chromatin. Certain TFs bind G4s with affinities comparable to double-stranded DNA targets. We demonstrate that, in a chromatin context, this binding interaction is competed out with a small molecule. Notably, endogenous G4s are prominent binding sites for a large number of TFs, particularly at promoters of highly expressed genes.
Conclusions: Our results reveal a novel non-canonical mechanism for TF binding whereby G4s operate as common binding hubs for many different TFs to promote increased transcription.
Keywords: Chemical biology; DNA G-quadruplex; Gene expression; Transcription factor binding.
Conflict of interest statement
S.B. is a founder and shareholder of Cambridge Epigenetix Ltd.
Figures
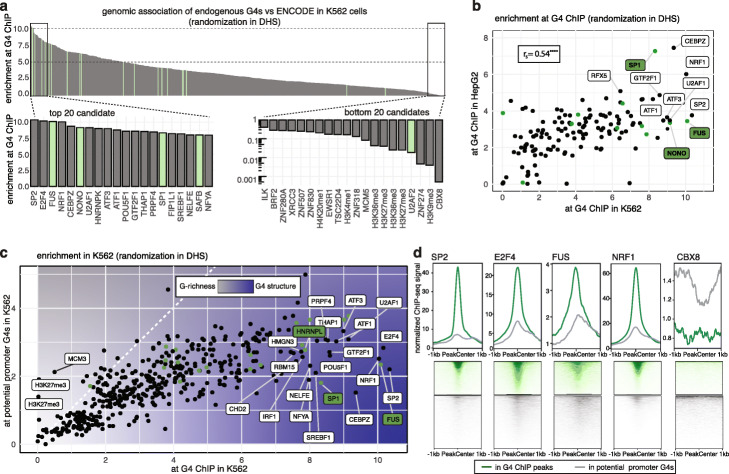
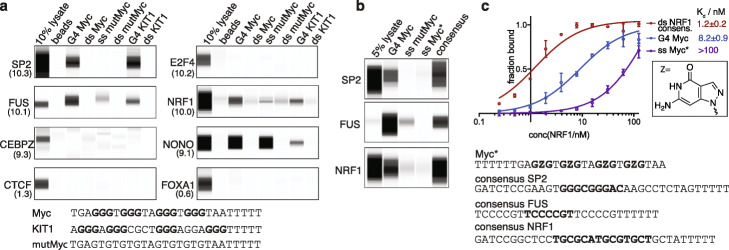
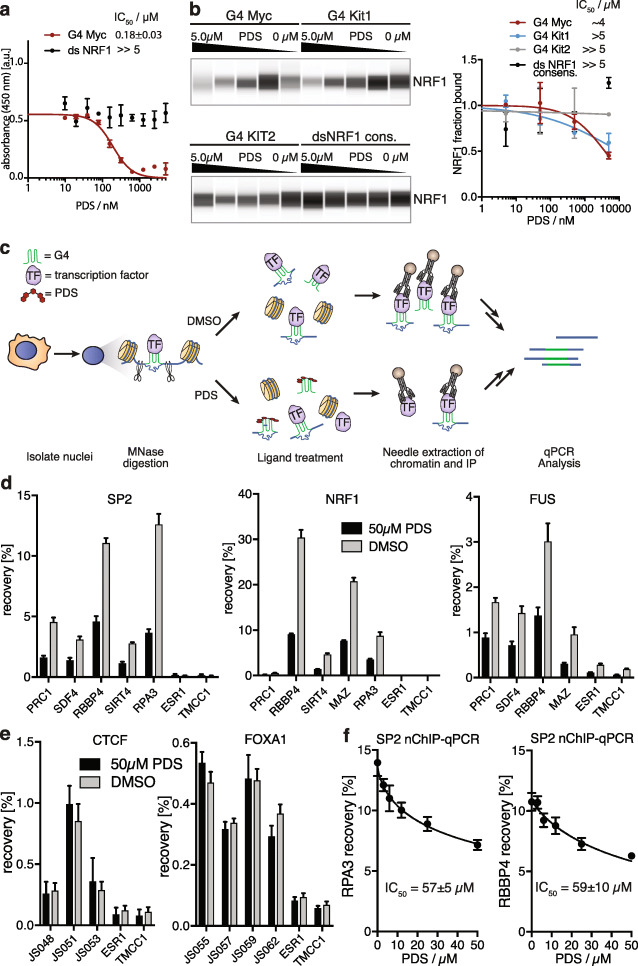
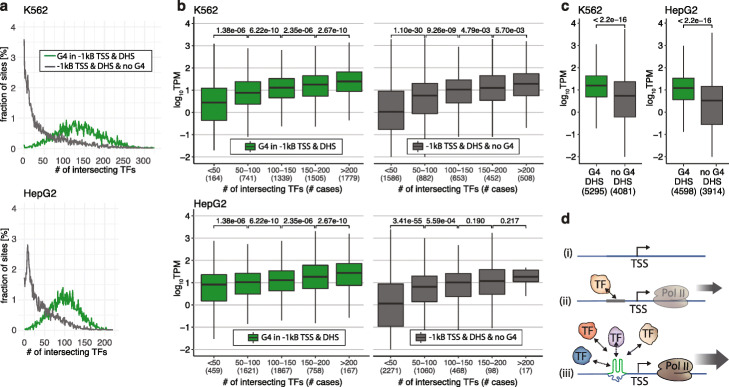
References
-
- Badis G, Berger MF, Philippakis AA, Talukder S, Gehrke AR, Jaeger SA, Chan ET, Metzler G, Vedenko A, Chen X, Kuznetsov H, Wang CF, Coburn D, Newburger DE, Morris Q, Hughes TR, Bulyk ML. Diversity and complexity in DNA recognition by transcription factors. Science. 2009;324(5935):1720–1723. doi: 10.1126/science.1162327. - DOI - PMC - PubMed
-
- Wang J, Zhuang J, Iyer S, Lin XY, Greven MC, Kim BH, et al. Factorbook.org: A Wiki-based database for transcription factor-binding data generated by the ENCODE consortium. Nucleic Acids Res. 2013;41:171–6. 10.1093/nar/gks1221. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
