

Robust induction of TRMs by combinatorial nanoshells confers cross-strain sterilizing immunity against lethal influenza viruses
- PMID: 33898629
- PMCID: PMC8047433
- DOI: 10.1016/j.omtm.2021.03.010
Robust induction of TRMs by combinatorial nanoshells confers cross-strain sterilizing immunity against lethal influenza viruses
Abstract
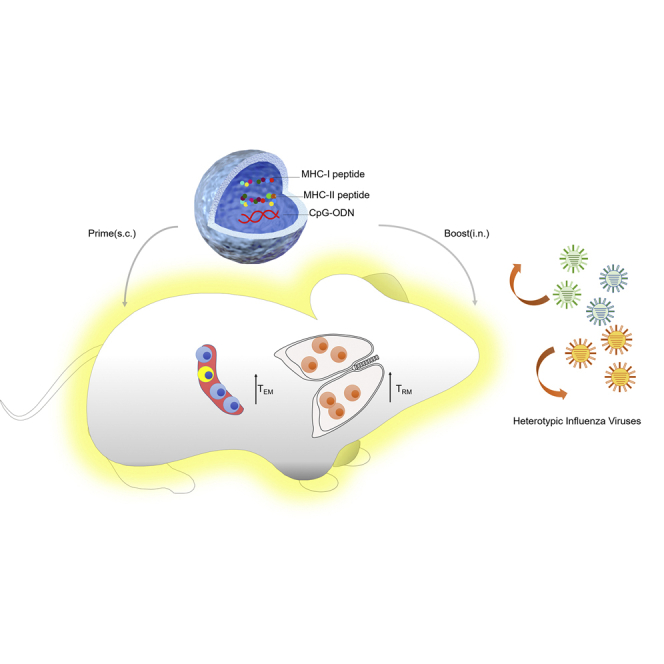
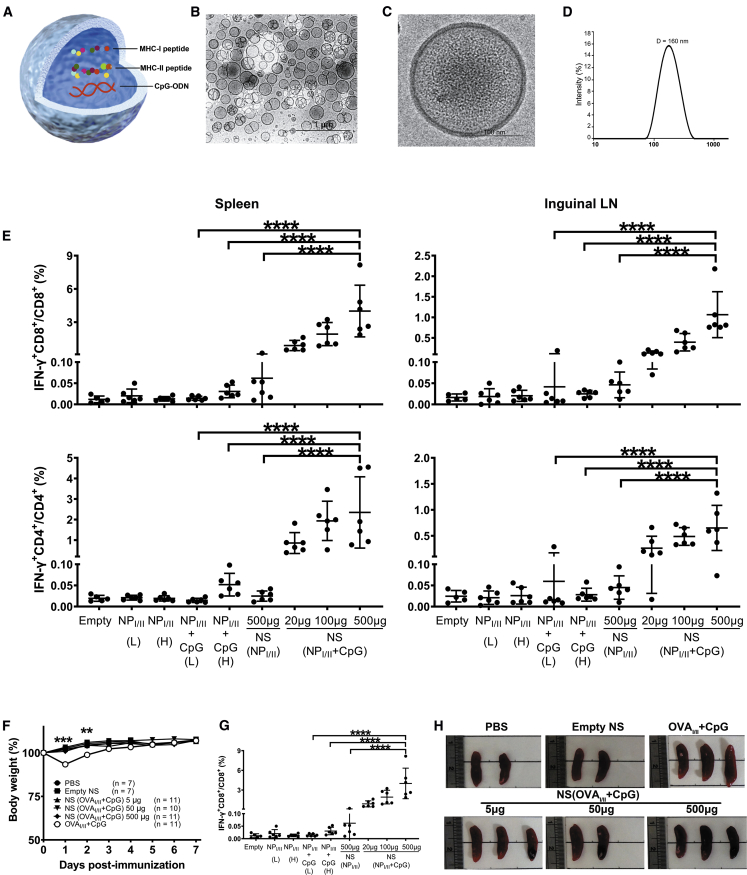
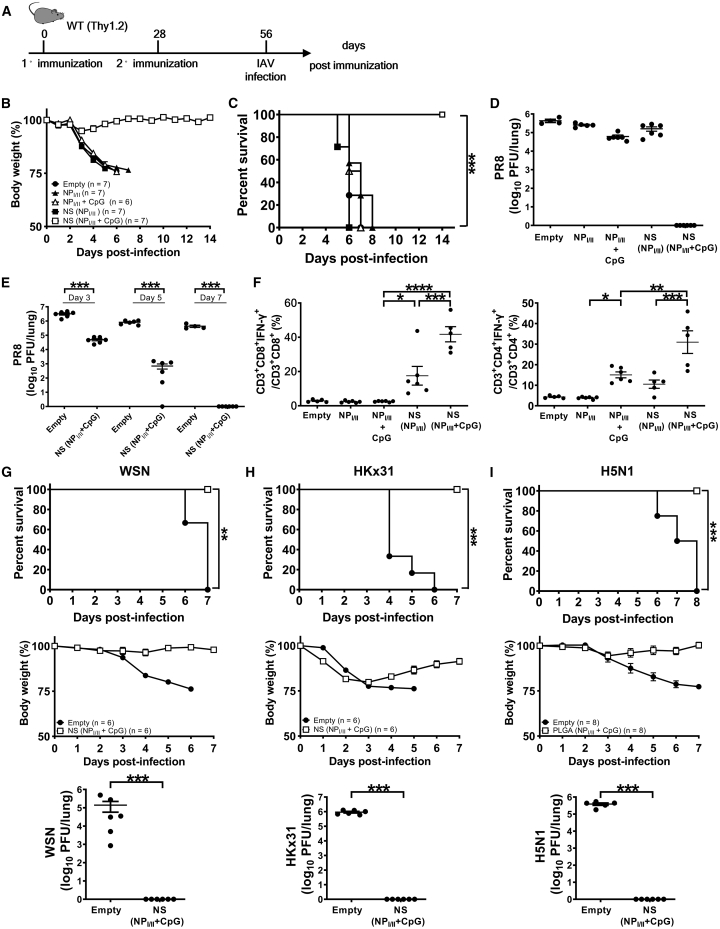
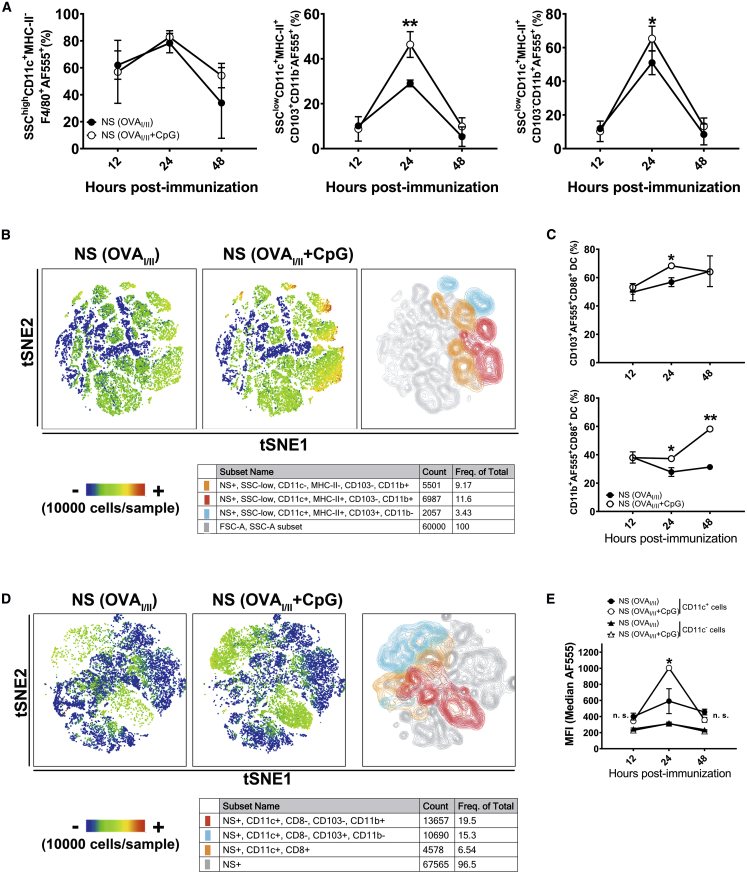
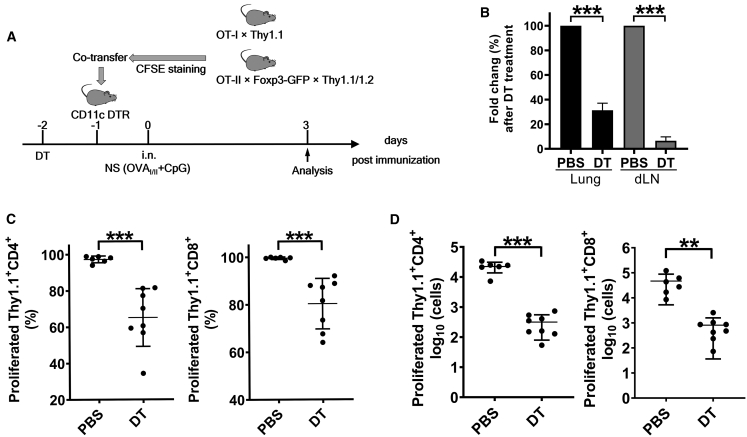
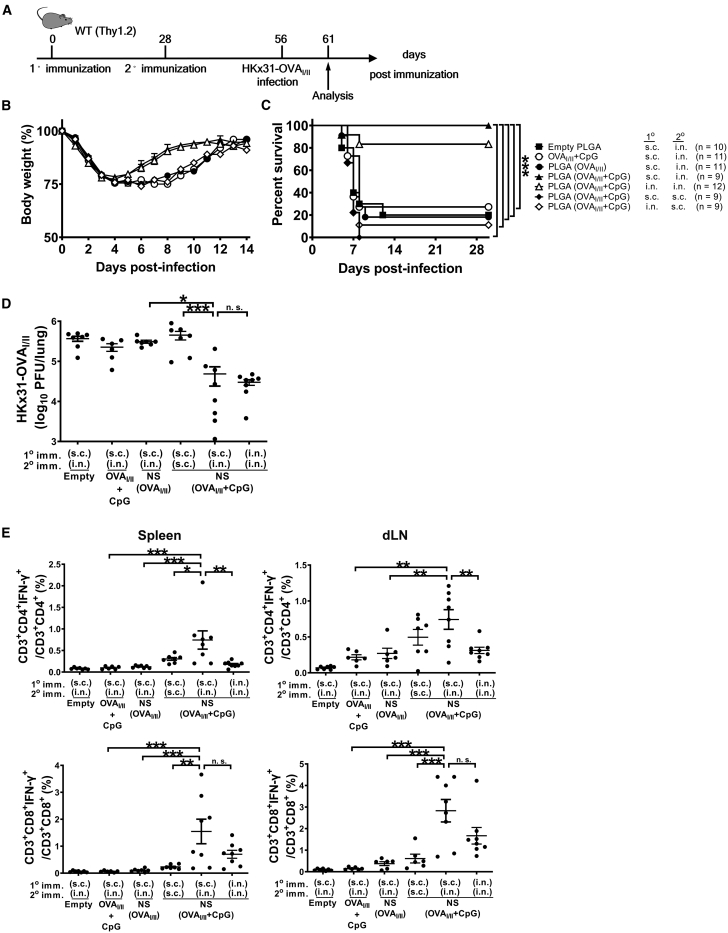
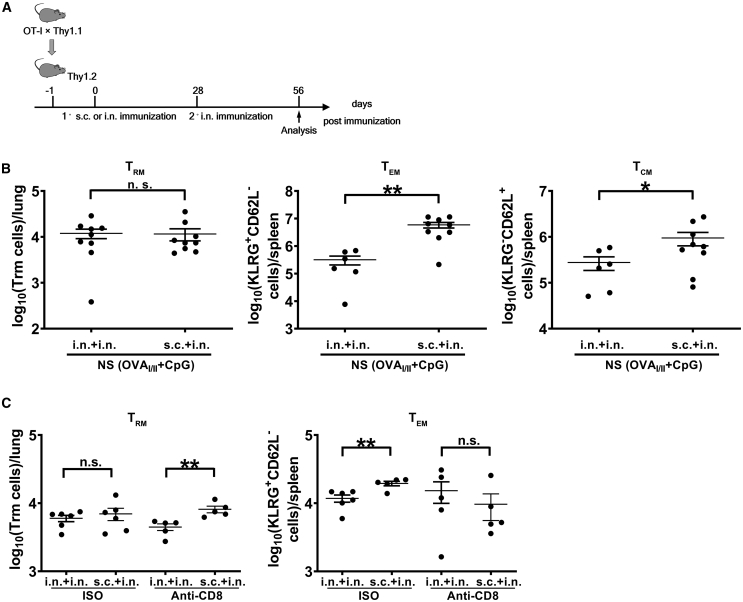
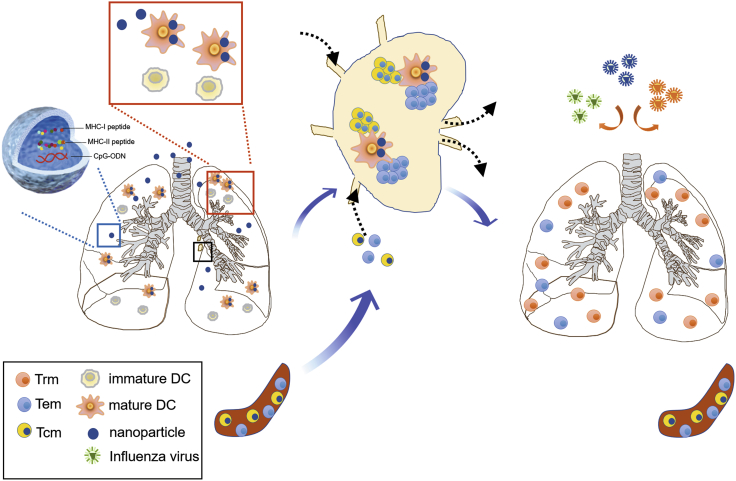
Antigen-specific lung-resident memory T cells (TRMs) constitute the first line of defense that mediates rapid protection against respiratory pathogens and inspires novel vaccine designs against infectious pandemic threats, yet effective means of inducing TRMs, particularly via non-viral vectors, remain challenging. Here, we demonstrate safe and potent induction of lung-resident TRMs using a biodegradable polymeric nanoshell that co-encapsulates antigenic peptides and TLR9 agonist CpG-oligodeoxynucleotide (CpG-ODN) in a virus-mimicking structure. Through subcutaneous priming and intranasal boosting, the combinatorial nanoshell vaccine elicits prominent lung-resident CD4+ and CD8+ T cells that surprisingly show better durability than live viral infections. In particular, nanoshells containing CpG-ODN and a pair of conserved class I and II major histocompatibility complex-restricted influenza nucleoprotein-derived antigenic peptides are demonstrated to induce near-sterilizing immunity against lethal infections with influenza A viruses of different strains and subtypes in mice, resulting in rapid elimination of replicating viruses. We further examine the pulmonary transport dynamic and optimal composition of the nanoshell vaccine conducive for robust TRM induction as well as the benefit of subcutaneous priming on TRM replenishment. The study presents a practical vaccination strategy for inducing protective TRM-mediated immunity, offering a compelling platform and critical insights in the ongoing quest toward a broadly protective vaccine against universal influenza as well as other respiratory pathogens.
Keywords: T cell vaccine; influenza virus; nanoshell; peptide vaccine; resident memory T cell; respiratory infection; universal influenza vaccine.
© 2021 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Increased Protein Degradation Improves Influenza Virus Nucleoprotein-Specific CD8+ T Cell Activation In Vitro but Not in C57BL/6 Mice.J Virol. 2016 Oct 28;90(22):10209-10219. doi: 10.1128/JVI.01633-16. Print 2016 Nov 15. J Virol. 2016. PMID: 27581985 Free PMC article.
-
A Role of Influenza Virus Exposure History in Determining Pandemic Susceptibility and CD8+ T Cell Responses.J Virol. 2016 Jul 11;90(15):6936-6947. doi: 10.1128/JVI.00349-16. Print 2016 Aug 1. J Virol. 2016. PMID: 27226365 Free PMC article.
-
CCR2 Regulates Vaccine-Induced Mucosal T-Cell Memory to Influenza A Virus.bioRxiv [Preprint]. 2021 Mar 25:2021.03.24.436901. doi: 10.1101/2021.03.24.436901. bioRxiv. 2021. Update in: J Virol. 2021 Jul 12;95(15):e0053021. doi: 10.1128/JVI.00530-21. PMID: 33791695 Free PMC article. Updated. Preprint.
-
Harnessing the Power of T Cells: The Promising Hope for a Universal Influenza Vaccine.Vaccines (Basel). 2018 Mar 26;6(2):18. doi: 10.3390/vaccines6020018. Vaccines (Basel). 2018. PMID: 29587436 Free PMC article. Review.
-
Progress on adenovirus-vectored universal influenza vaccines.Hum Vaccin Immunother. 2015;11(5):1209-22. doi: 10.1080/21645515.2015.1016674. Hum Vaccin Immunother. 2015. PMID: 25876176 Free PMC article. Review.
Cited by
-
Lymph Node Follicle-Targeting STING Agonist Nanoshells Enable Single-Shot M2e Vaccination for Broad and Durable Influenza Protection.Adv Sci (Weinh). 2023 Jun;10(17):e2206521. doi: 10.1002/advs.202206521. Epub 2023 Apr 24. Adv Sci (Weinh). 2023. PMID: 37092580 Free PMC article.
-
A molecular signature of lung-resident CD8+ T cells elicited by subunit vaccination.Sci Rep. 2022 Nov 9;12(1):19101. doi: 10.1038/s41598-022-21620-7. Sci Rep. 2022. PMID: 36351985 Free PMC article.
-
Tissue resident memory T cells in the respiratory tract.Mucosal Immunol. 2022 Mar;15(3):379-388. doi: 10.1038/s41385-021-00461-z. Epub 2021 Oct 20. Mucosal Immunol. 2022. PMID: 34671115 Free PMC article. Review.
-
Influenza Virus-Derived CD8 T Cell Epitopes: Implications for the Development of Universal Influenza Vaccines.Immune Netw. 2024 May 7;24(3):e19. doi: 10.4110/in.2024.24.e19. eCollection 2024 Jun. Immune Netw. 2024. PMID: 38974213 Free PMC article. Review.
-
Intranasal Epitope-Polymer Vaccine Lodges Resident Memory T Cells Protecting Against Influenza Virus.Adv Healthc Mater. 2024 Jun;13(15):e2304188. doi: 10.1002/adhm.202304188. Epub 2024 Mar 9. Adv Healthc Mater. 2024. PMID: 38411375 Free PMC article.
References
-
- Mueller S.N., Mackay L.K. Tissue-resident memory T cells: local specialists in immune defence. Nat. Rev. Immunol. 2016;16:79–89. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
