

EZH2 inhibition activates a dsRNA-STING-interferon stress axis that potentiates response to PD-1 checkpoint blockade in prostate cancer
- PMID: 33899001
- PMCID: PMC8061902
- DOI: 10.1038/s43018-021-00185-w
EZH2 inhibition activates a dsRNA-STING-interferon stress axis that potentiates response to PD-1 checkpoint blockade in prostate cancer
Abstract
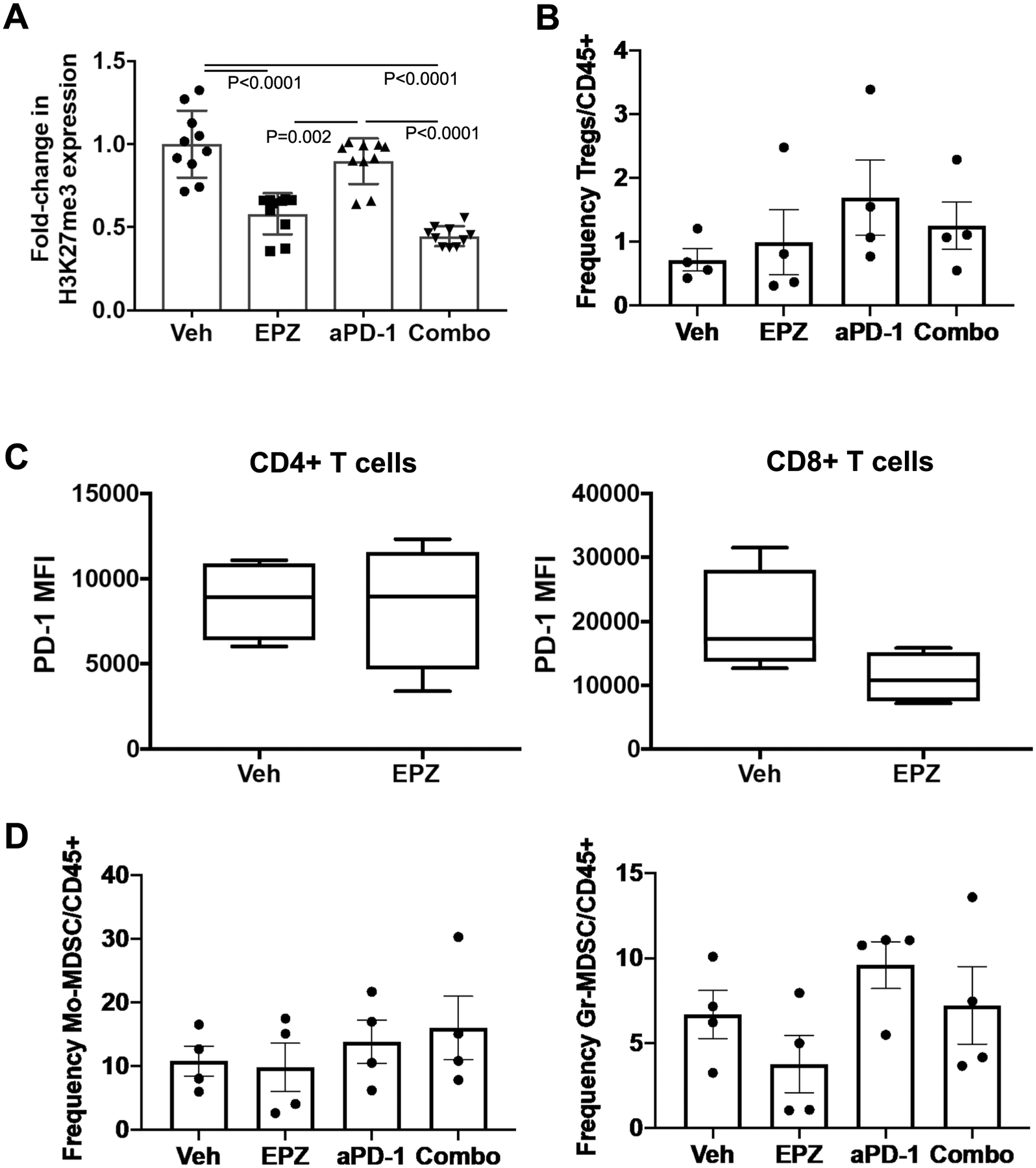
Prostate cancers are considered to be immunologically 'cold' tumors given the very few patients who respond to checkpoint inhibitor (CPI) therapy. Recently, enrichment of interferon-stimulated genes (ISGs) predicted a favorable response to CPI across various disease sites. The enhancer of zeste homolog-2 (EZH2) is overexpressed in prostate cancer and known to negatively regulate ISGs. In the present study, we demonstrate that EZH2 inhibition in prostate cancer models activates a double-stranded RNA-STING-ISG stress response upregulating genes involved in antigen presentation, Th1 chemokine signaling and interferon response, including programmed cell death protein 1 (PD-L1) that is dependent on STING activation. EZH2 inhibition substantially increased intratumoral trafficking of activated CD8+ T cells and increased M1 tumor-associated macrophages, overall reversing resistance to PD-1 CPI. Our study identifies EZH2 as a potent inhibitor of antitumor immunity and responsiveness to CPI. These data suggest EZH2 inhibition as a therapeutic direction to enhance prostate cancer response to PD-1 CPI.
Conflict of interest statement
Competing interests The authors declare no competing interests.
Figures













References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
