

Shikimic acid protects skin cells from UV-induced senescence through activation of the NAD+-dependent deacetylase SIRT1
- PMID: 33901008
- PMCID: PMC8148468
- DOI: 10.18632/aging.203010
Shikimic acid protects skin cells from UV-induced senescence through activation of the NAD+-dependent deacetylase SIRT1
Abstract
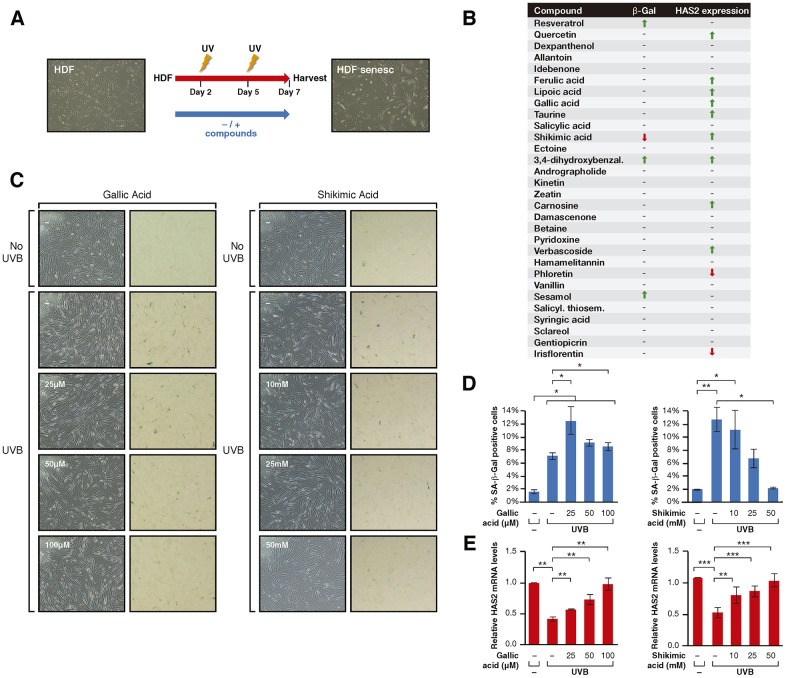
UV radiation is one of the main contributors to skin photoaging by promoting the accumulation of cellular senescence, which in turn induces a proinflammatory and tissue-degrading state that favors skin aging. The members of the sirtuin family of NAD+-dependent enzymes play an anti-senescence role and their activation suggests a promising approach for preventing UV-induced senescence in the treatment of skin aging. A two-step screening designed to identify compounds able to protect cells from UV-induced senescence through sirtuin activation identified shikimic acid (SA), a metabolic intermediate in many organisms, as a bona-fide candidate. The protective effects of SA against senescence were dependent on specific activation of SIRT1 as the effect was abrogated by the SIRT1 inhibitor EX-527. Upon UV irradiation SA induced S-phase accumulation and a decrease in p16INK4A expression but did not protect against DNA damage or increased polyploidies. In contrast, SA reverted misfolded protein accumulation upon senescence, an effect that was abrogated by EX-527. Consistently, SA induced an increase in the levels of the chaperone BiP, resulting in a downregulation of unfolded protein response (UPR) signaling and UPR-dependent autophagy, avoiding their abnormal hyperactivation during senescence. SA did not directly activate SIRT1 in vitro, suggesting that SIRT1 is a downstream effector of SA signaling specifically in the response to cellular senescence. Our study not only uncovers a shikimic acid/SIRT1 signaling pathway that prevents cellular senescence, but also reinforces the role of sirtuins as key regulators of cell proteostasis.
Keywords: SIRT1; UV irradiation; human dermal fibroblasts; senescence; shikimic acid.
Conflict of interest statement
Figures




References
-
- Sjerobabski-Masnec I, Situm M. Skin aging. Acta Clin Croat. 2010; 49:515–18. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
