

The Stochastic Genome and Its Role in Gene Expression
- PMID: 33903158
- PMCID: PMC8485747
- DOI: 10.1101/cshperspect.a040386
The Stochastic Genome and Its Role in Gene Expression
Abstract
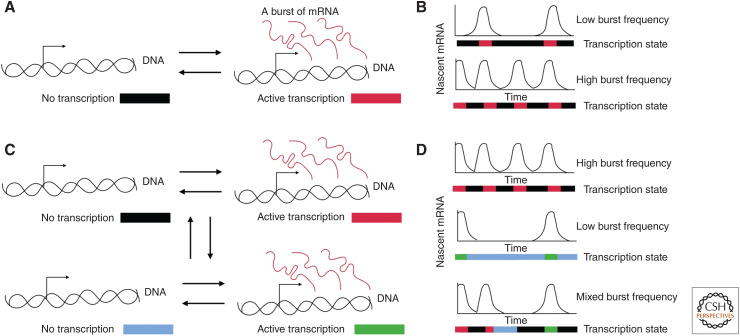
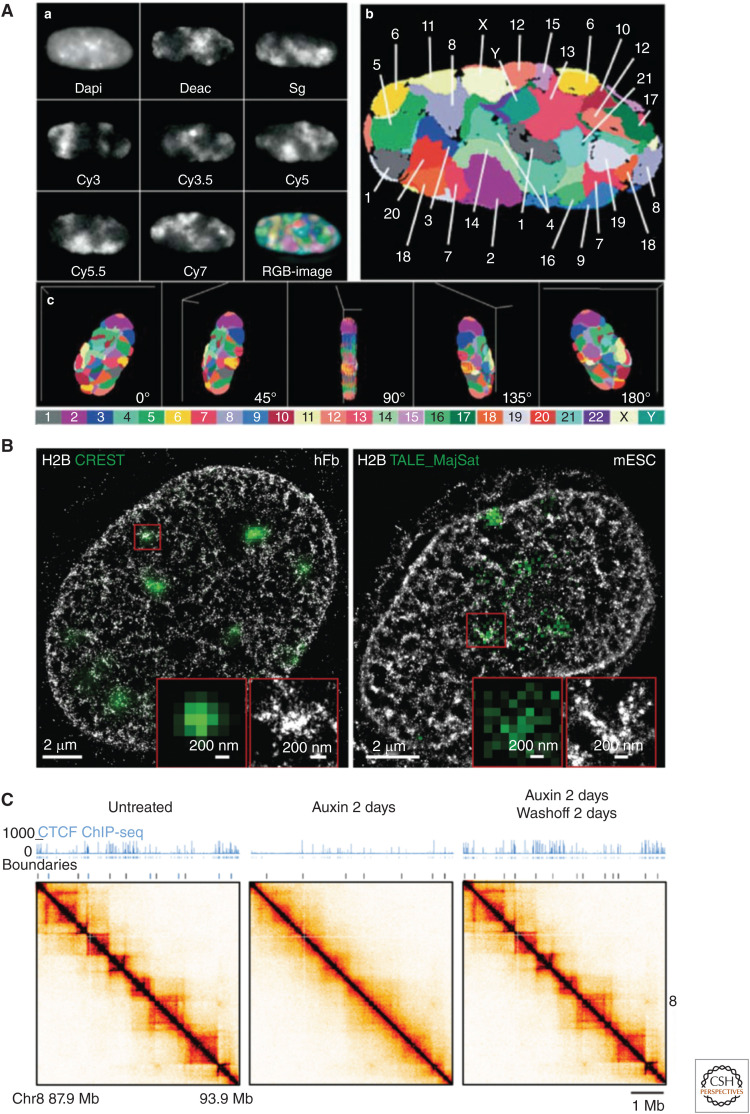
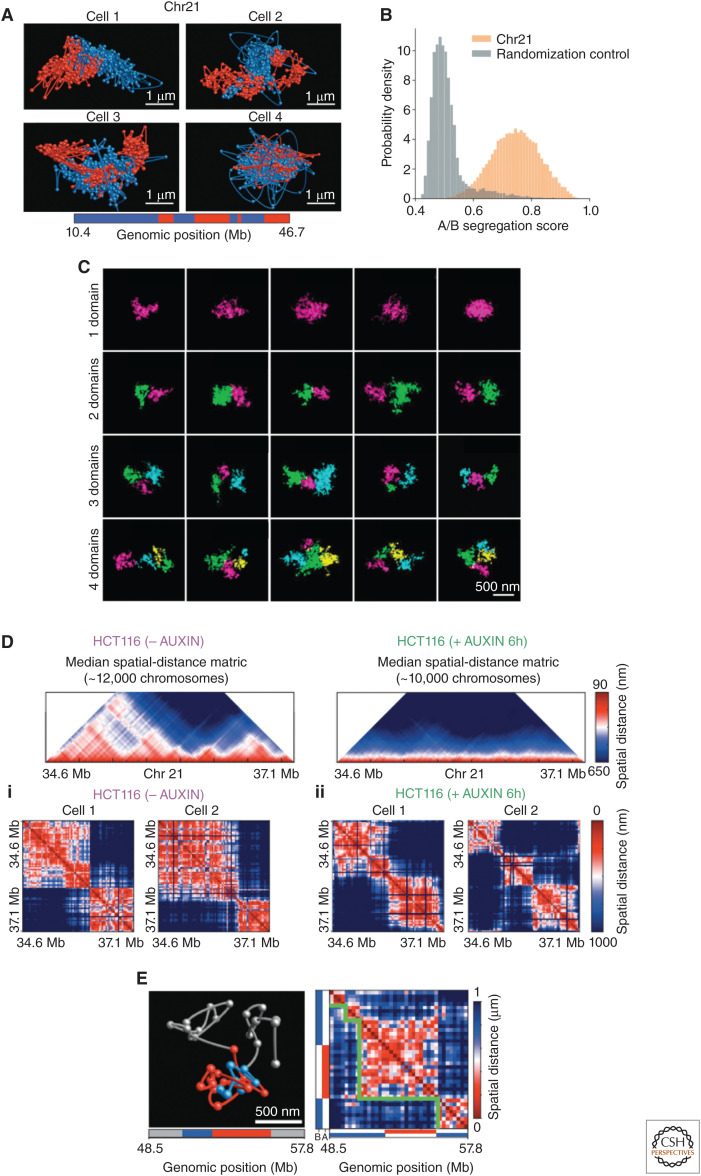
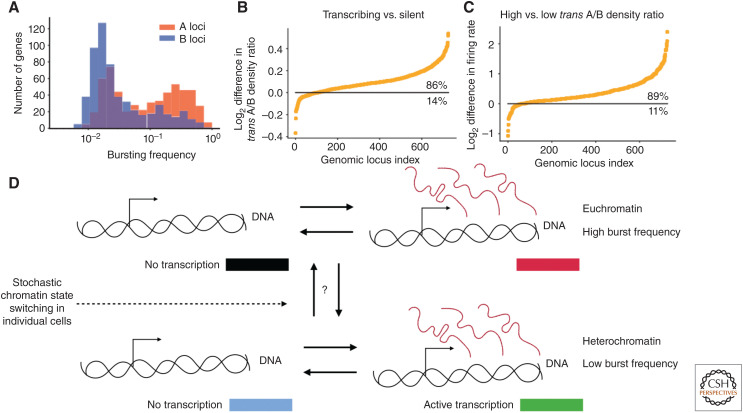
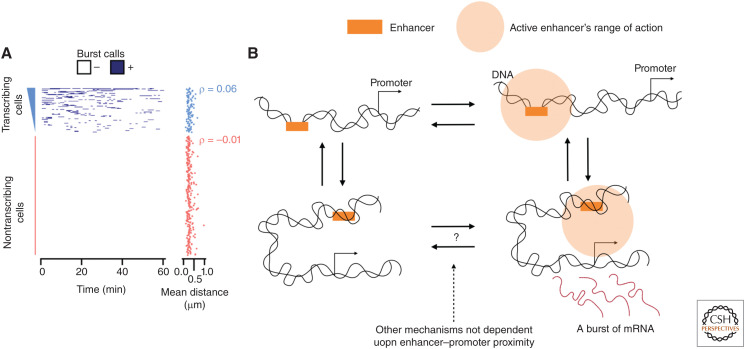
Mammalian genomes have distinct levels of spatial organization and structure that have been hypothesized to play important roles in transcription regulation. Although much has been learned about these architectural features with ensemble techniques, single-cell studies are showing a new universal trend: Genomes are stochastic and dynamic at every level of organization. Stochastic gene expression, on the other hand, has been studied for years. In this review, we probe whether there is a causative link between the two phenomena. We specifically discuss the functionality of chromatin state, topologically associating domains (TADs), and enhancer biology in light of their stochastic nature and their specific roles in stochastic gene expression. We highlight persistent fundamental questions in this area of research.
Copyright © 2021 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures

References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources