

Review
doi: 10.1038/s41556-021-00669-y.
Epub 2021 Apr 26.
Autophagosome biogenesis comes out of the black box
Affiliations
- PMID: 33903736
- PMCID: PMC8122082
- DOI: 10.1038/s41556-021-00669-y
Item in Clipboard
Review
Autophagosome biogenesis comes out of the black box
Nat Cell Biol.
2021 May.
Abstract
Macroautophagic clearance of cytosolic materials entails the initiation, growth and closure of autophagosomes. Cargo triggers the assembly of a web of cargo receptors and core machinery. Autophagy-related protein 9 (ATG9) vesicles seed the growing autophagosomal membrane, which is supplied by de novo phospholipid synthesis, phospholipid transport via ATG2 proteins and lipid flipping by ATG9. Autophagosomes close via ESCRT complexes. Here, we review recent discoveries that illuminate the molecular mechanisms of autophagosome formation and discuss emerging questions in this rapidly developing field.
Conflict of interest statement
Conflicts of interest
J.H.H. is a co-founder of Casma Therapeutics.
Figures
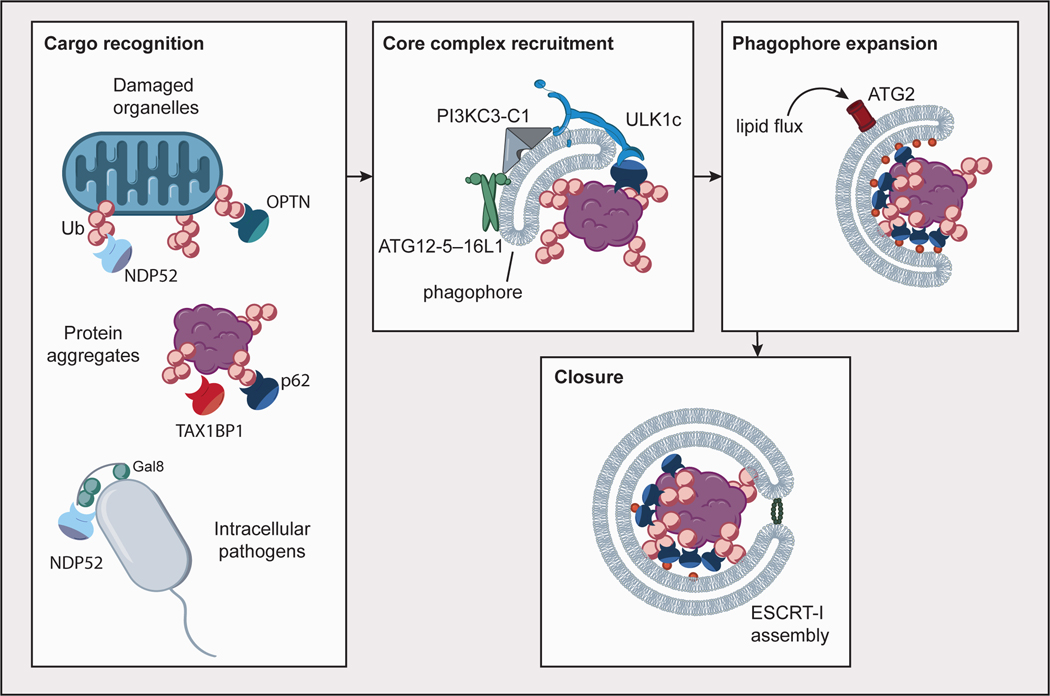
Autophagy cargoes such as damaged organelles, protein aggregates and intracellular pathogens are recognized by cargo receptors such as p62, NDP52, TAX1BP1, and OPTN, most commonly through modification with ubiquitin (Ub), as well as the presence of other signals such as Galectin-8. Core autophagy complexes (ULK1c, PI3KC3-C1, ATG12–ATG5–16L1) are recruited to the nascent phagophore through interactions with cargo receptors, the phagophore membrane, and one another in a positive feedback mechanism. Phagophore expansion is driven by lipid flux through ATG2. Autophagosome closure is mediated in part by assembly of ESCRT-I proteins in a ring at the aperture of the closing phagophore.
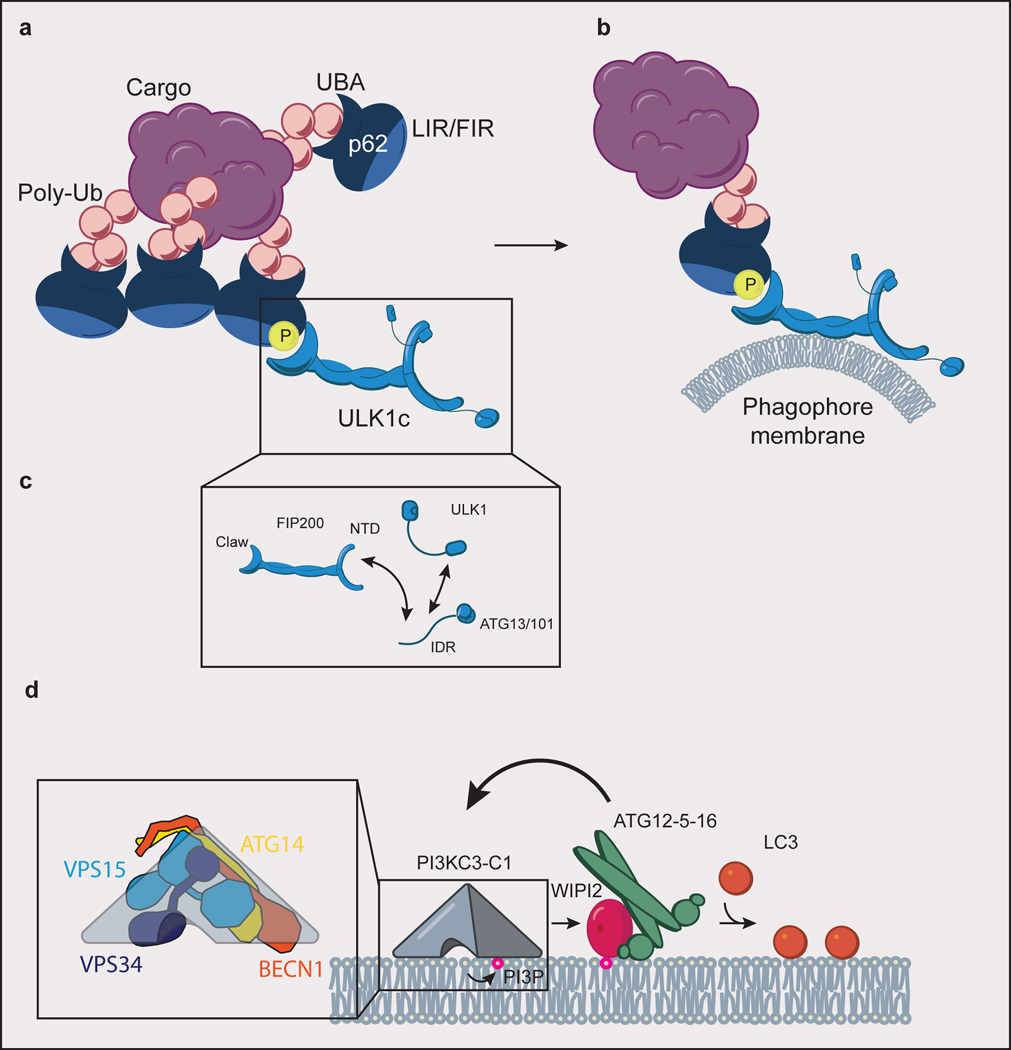
(a) ULK1c is recruited to p62 condensate cargo through an interaction of the FIP200 claw domain with phospho-p62. p62 binds to ubiquitinated (Ub) cargo via its UBA domain, to LC3 via its LIR motif, and to FIP200 of the ULK1c via its FIR motif. (b) Interaction of ULK1c with p62 condensates stimulates “on demand” phagophore membrane generation. (c) The subunits of ULK1c (FIP200, ULK1, ATG13, ATG13/101) are held together in large part through a network of interacting IDRs. (d) PI3KC3-C1 (inset: comprised of ATG14, BECN1, VPS15, and VPS34) phosphorylates phosphatidylinositol lipid headgroups to form PI(3)P, which recruits WIPI proteins to the phagophore membrane. WIPI2 subsequently recruits ATG12–5–16 to the phagophore in a positive feedback loop with PI3KC3-C1. ATG12–5–16 covalently links LC3 to phosphatidylethanolamine lipid headgroups (orange circles) in the phagophore membrane.
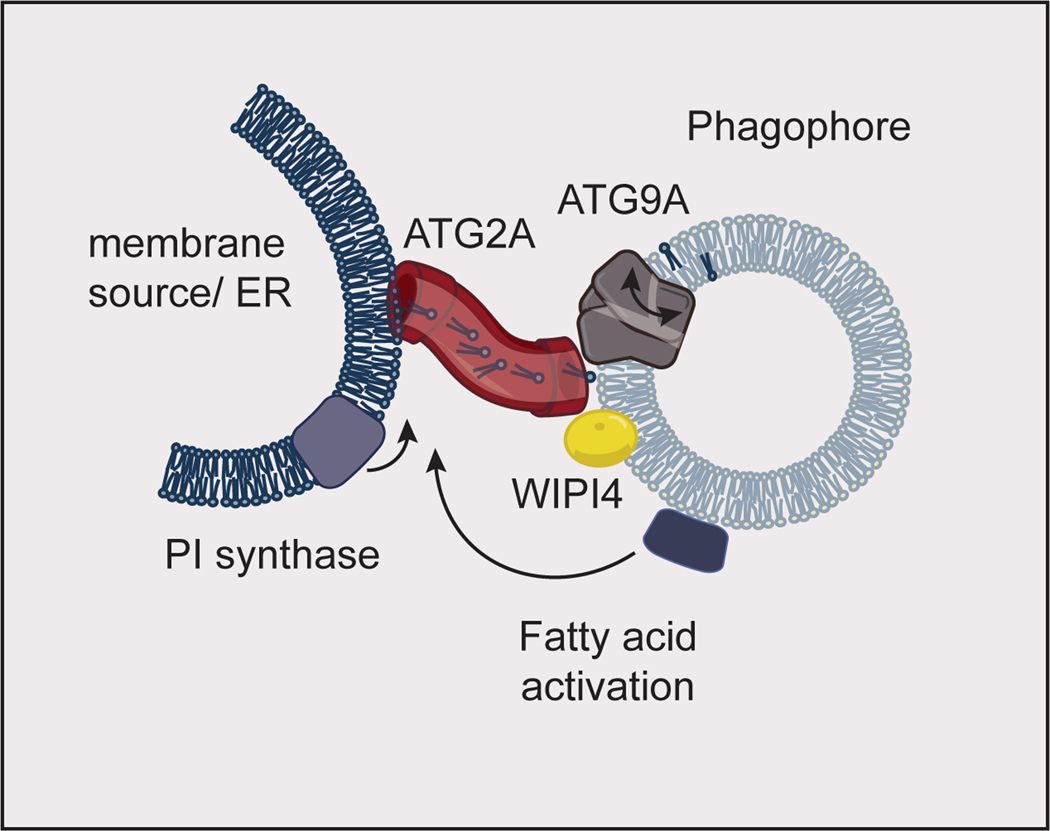
Fatty acid activation at the phagophore feeds phospholipid synthesis by phosphatidylinositol (PI) synthase at the membrane source for the autophagosome. This phospholipid synthesis drives lipid flux through ATG2A (which binds WIPI4 on the phagophore) into the growing phagophore membrane. ATG9A aids in phagophore growth by scrambling lipids between the inner and outer leaflets of the phagophore. The model shown here is inspired by data obtained in yeast. The proteins implicated in autophagosome growth are conserved between yeast and humans, and human proteins are shown here. ER: Endoplasmic Reticulum.
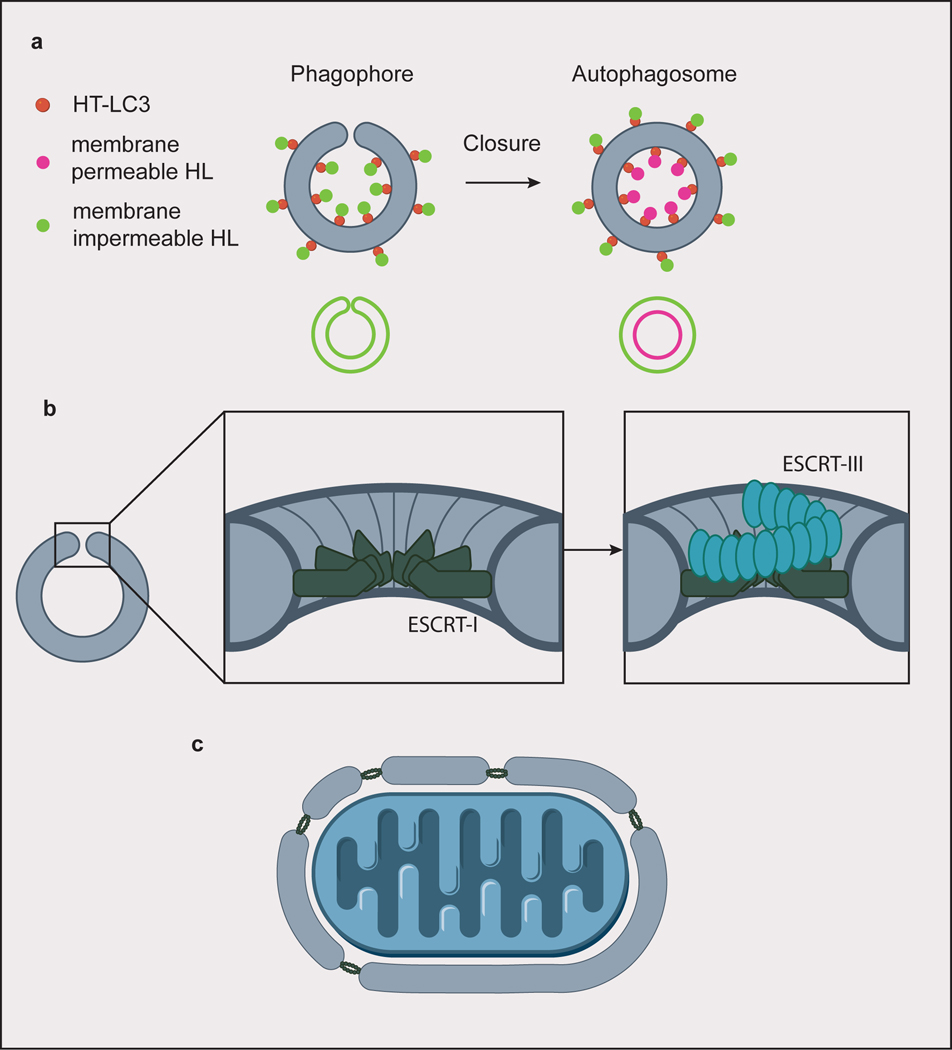
(a) A screen for factors involved in autophagosome closure identified ESCRT-I protein VPS37A by using membrane permeable/impermeable Halo ligand (HL) dyes to selectively label Halo tagged (HT) LC3. Hits were identified from cells whose phagophores only stained with the impermeable dye, indicating failure to close the autophagosomes. Closed autophagosomes stain with both the membrane permeable and impermeable dyes. (b) ESCRT-I forms a dodecameric ring assembly at the rim of the closing autophagosome. In canonical ESCRT-mediated membrane scission, subsequent helical assembly of ESCRT-III constricts the membrane rim. (c) For larger cargoes, ESCRTs could be involved in piecing together membranes to form a single, large phagophore.
References
-
- Mizushima N, Yoshimori T & Ohsumi Y. The role of Atg proteins in autophagosome formation. Annu Rev Cell Dev Biol 27, 107–132 (2011). - PubMed
-
- Lamb CA, Yoshimori T & Tooze SA The autophagosome: origins unknown, biogenesis complex. Nature Reviews Molecular Cell Biology 14, 759 (2013). - PubMed
-
- Nakatogawa H. Mechanisms governing autophagosome biogenesis. Nat Rev Mol Cell Biol 21, 439–458 (2020). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
