

Mixed xenogeneic porcine chimerism tolerizes human anti-pig natural antibody-producing cells in a humanized mouse model
- PMID: 33904221
- PMCID: PMC8376778
- DOI: 10.1111/xen.12691
Mixed xenogeneic porcine chimerism tolerizes human anti-pig natural antibody-producing cells in a humanized mouse model
Abstract
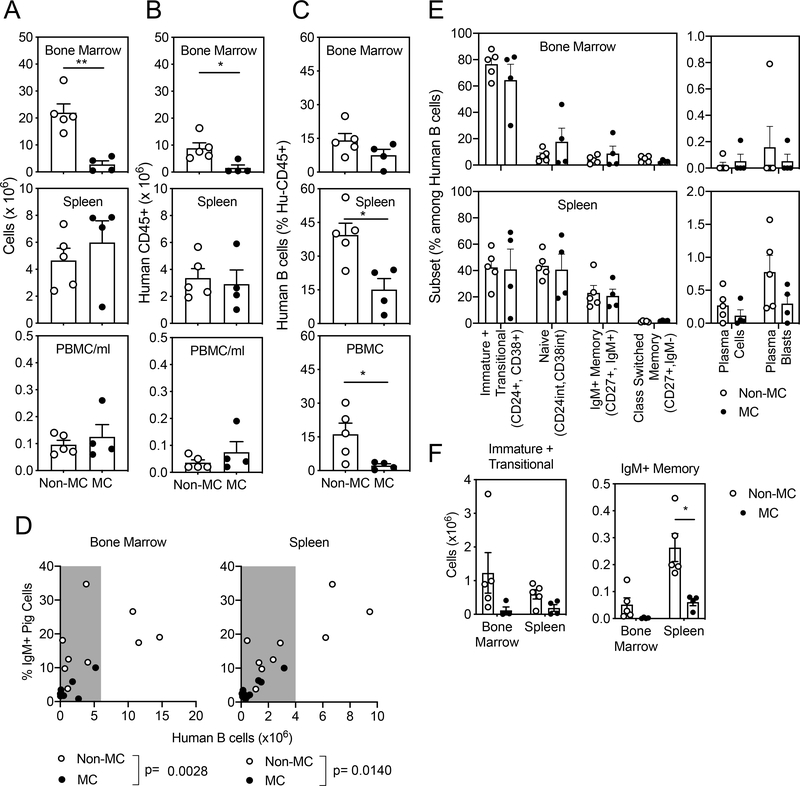
Background: A major obstacle to the success of organ transplantation from pigs to humans, necessitated by the shortage of human organs, is robust humoral immune rejection by pig-reactive human antibodies. Mixed xenogeneic hematopoietic chimerism induces xenoreactive B cell tolerance in rodents, but whether mixed pig/human chimerism could induce tolerance of human B cells to pig xenoantigens is unknown.
Methods: We investigated this question using a humanized mouse model in which durable mixed (pig-human) xenogeneic chimerism can be established.
Results: Human natural anti-pig cytotoxic antibodies, predominantly IgM, are detectable in non-chimeric humanized mouse serum, and pig-reactive antibodies were reduced in mixed chimeric versus non-chimeric humanized mice. This difference required persistent mixed chimerism and was not due to the adsorption of antibodies on pig cells in vivo. Furthermore, human B cells from spleens of mixed chimeric mice produced lower levels of anti-pig antibodies when stimulated in vitro compared with those from non-chimeric mice.
Conclusions: Our findings demonstrate that mixed chimerism reduces human natural antibodies to pig xenoantigens, providing the first in vivo evidence of human B cell tolerance induction by mixed xenogeneic chimerism and supporting further evaluation of this approach for inducing human B cell tolerance to xenografts.
Keywords: B cells; Mixed Chimerism; humanized mice; xenoantibody; xenotolerance.
© 2021 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd.
Conflict of interest statement
Figures






References
-
- Galili U Anti-Gal in Humans and Its Antigen the α-Gal Epitope. In: The Natural Anti-Gal Antibody As Foe Turned Friend In Medicine.2018:3–22.
-
- Lai L, Kolber-Simonds D, Park KW, et al.Production of alpha-1,3-galactosyltransferase knockout pigs by nuclear transfer cloning. Science. 2002;295(5557):1089–1092. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
