

Ubiquitin-proteasome system and the role of its inhibitors in cancer therapy
- PMID: 33906413
- PMCID: PMC8080017
- DOI: 10.1098/rsob.200390
Ubiquitin-proteasome system and the role of its inhibitors in cancer therapy
Abstract
Despite all the other cells that have the potential to prevent cancer development and metastasis through tumour suppressor proteins, cancer cells can upregulate the ubiquitin-proteasome system (UPS) by which they can degrade tumour suppressor proteins and avoid apoptosis. This system plays an extensive role in cell regulation organized in two steps. Each step has an important role in controlling cancer. This demonstrates the importance of understanding UPS inhibitors and improving these inhibitors to foster a new hope in cancer therapy. UPS inhibitors, as less invasive chemotherapy drugs, are increasingly used to alleviate symptoms of various cancers in malignant states. Despite their success in reducing the development of cancer with the lowest side effects, thus far, an appropriate inhibitor that can effectively inactivate this system with the least drug resistance has not yet been fully investigated. A fundamental understanding of the system is necessary to fully elucidate its role in causing/controlling cancer. In this review, we first comprehensively investigate this system, and then each step containing ubiquitination and protein degradation as well as their inhibitors are discussed. Ultimately, its advantages and disadvantages and some perspectives for improving the efficiency of these inhibitors are discussed.
Keywords: cancer; protein degradation inhibitors; targeted therapy; ubiquitination inhibitors; ubiquitin–proteasome system.
Figures
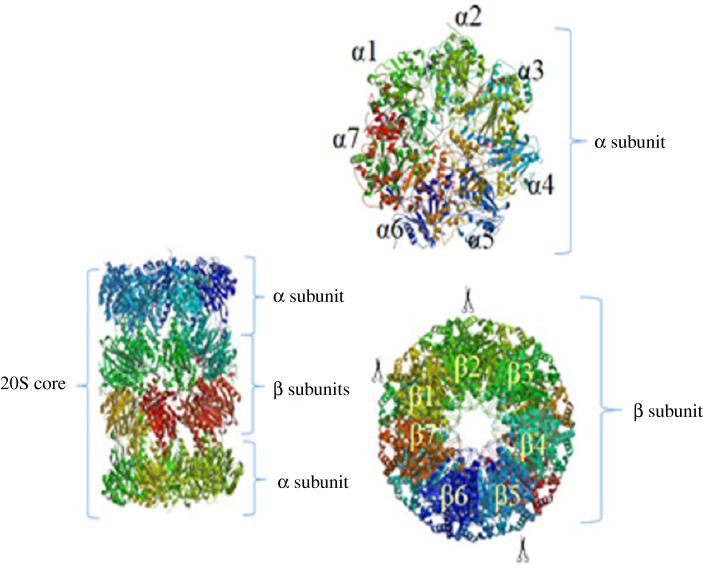
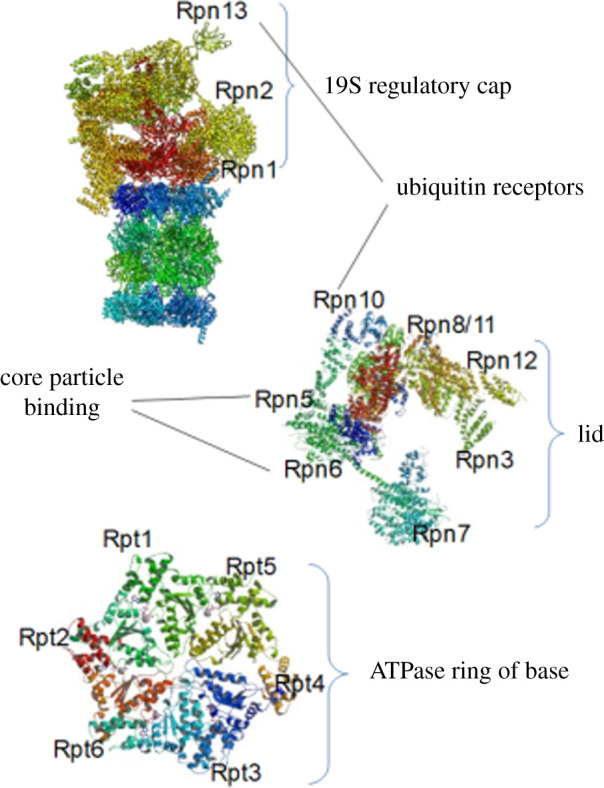
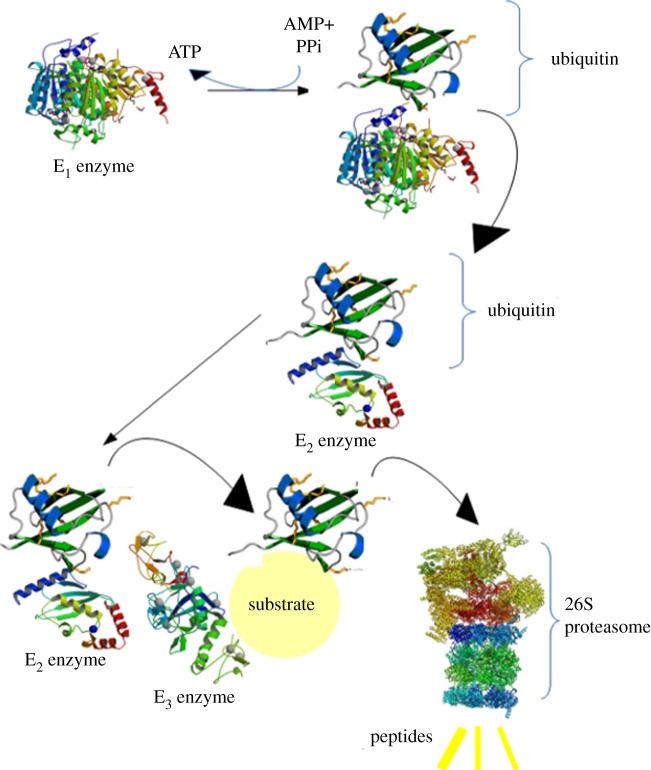
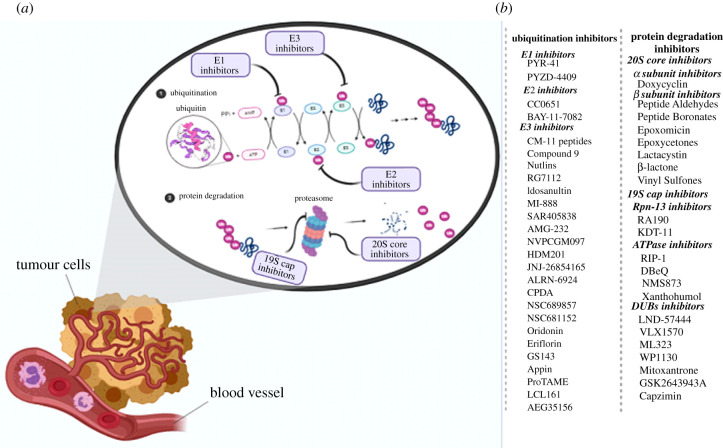
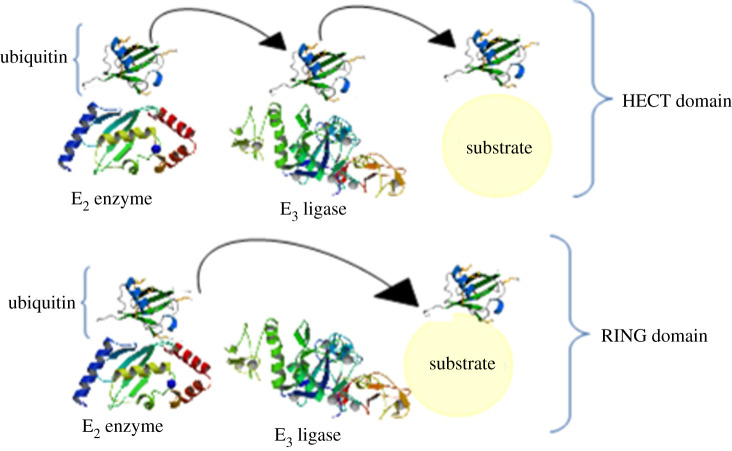
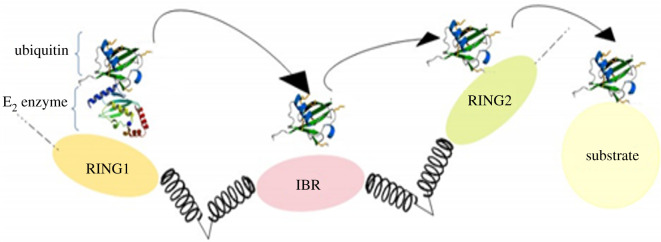
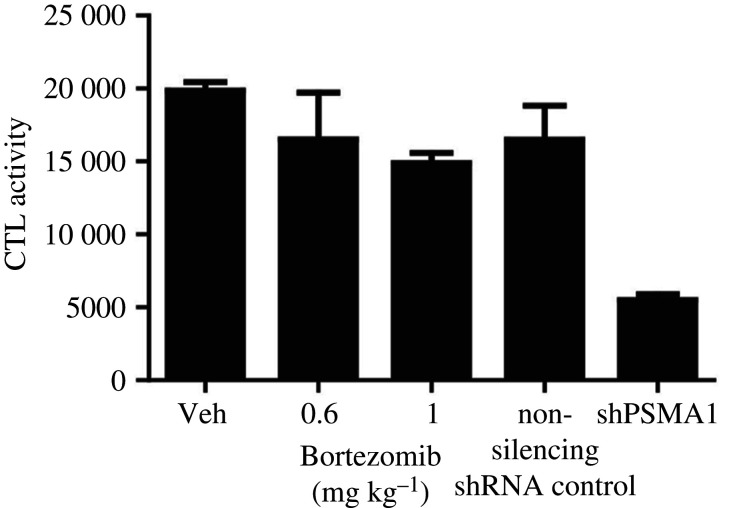
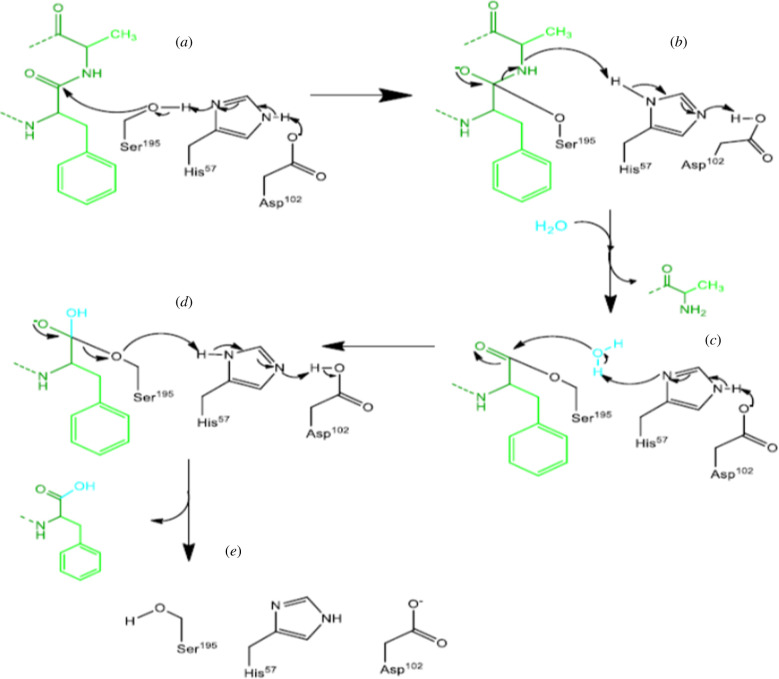
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical