

Anaerobic Microbial Metabolism of Dichloroacetate
- PMID: 33906923
- PMCID: PMC8092247
- DOI: 10.1128/mBio.00537-21
Anaerobic Microbial Metabolism of Dichloroacetate
Abstract
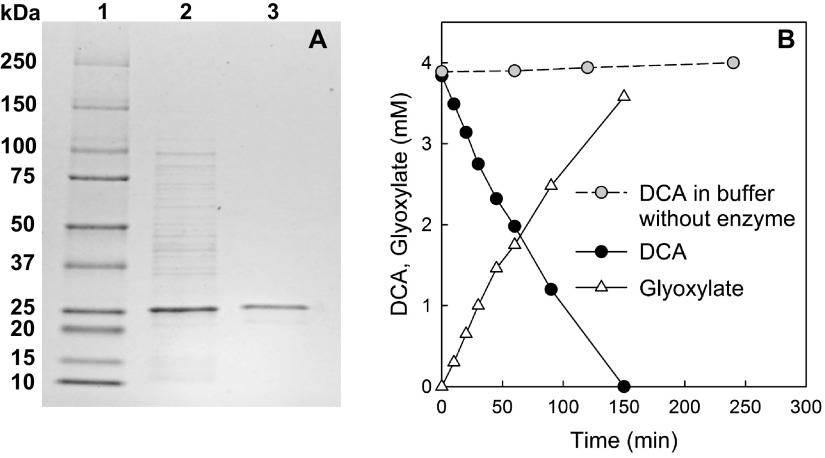
Dichloroacetate (DCA) commonly occurs in the environment due to natural production and anthropogenic releases, but its fate under anoxic conditions is uncertain. Mixed culture RM comprising "Candidatus Dichloromethanomonas elyunquensis" strain RM utilizes DCA as an energy source, and the transient formation of formate, H2, and carbon monoxide (CO) was observed during growth. Only about half of the DCA was recovered as acetate, suggesting a fermentative catabolic route rather than a reductive dechlorination pathway. Sequencing of 16S rRNA gene amplicons and 16S rRNA gene-targeted quantitative real-time PCR (qPCR) implicated "Candidatus Dichloromethanomonas elyunquensis" strain RM in DCA degradation. An (S)-2-haloacid dehalogenase (HAD) encoded on the genome of strain RM was heterologously expressed, and the purified HAD demonstrated the cofactor-independent stoichiometric conversion of DCA to glyoxylate at a rate of 90 ± 4.6 nkat mg-1 protein. Differential protein expression analysis identified enzymes catalyzing the conversion of DCA to acetyl coenzyme A (acetyl-CoA) via glyoxylate as well as enzymes of the Wood-Ljungdahl pathway. Glyoxylate carboligase, which catalyzes the condensation of two molecules of glyoxylate to form tartronate semialdehyde, was highly abundant in DCA-grown cells. The physiological, biochemical, and proteogenomic data demonstrate the involvement of an HAD and the Wood-Ljungdahl pathway in the anaerobic fermentation of DCA, which has implications for DCA turnover in natural and engineered environments, as well as the metabolism of the cancer drug DCA by gut microbiota.IMPORTANCE Dichloroacetate (DCA) is ubiquitous in the environment due to natural formation via biological and abiotic chlorination processes and the turnover of chlorinated organic materials (e.g., humic substances). Additional sources include DCA usage as a chemical feedstock and cancer drug and its unintentional formation during drinking water disinfection by chlorination. Despite the ubiquitous presence of DCA, its fate under anoxic conditions has remained obscure. We discovered an anaerobic bacterium capable of metabolizing DCA, identified the enzyme responsible for DCA dehalogenation, and elucidated a novel DCA fermentation pathway. The findings have implications for the turnover of DCA and the carbon and electron flow in electron acceptor-depleted environments and the human gastrointestinal tract.
Keywords: anaerobic catabolic pathways; comparative proteomics; dichloroacetate; fermentation; haloacid dehalogenase.
Copyright © 2021 Chen et al.
Figures





References
-
- Gribble GW. 2010. Progress in the chemistry of organic natural products, vol 91, p 12–13. Springer, Vienna, Austria.
-
- Matucha M, Gryndler M, Forczek ST, Uhlířová H, Fuksová K, Schröder P. 2003. Chloroacetic acids in environmental processes. Environ Chem Lett 1:127–130. doi: 10.1007/s10311-003-0030-y. - DOI
-
- McConnell O, Fenical W. 1977. Halogen chemistry of the red alga Asparagopsis. Phytochemistry 16:367–374. doi: 10.1016/0031-9422(77)80067-8. - DOI
-
- Moore RE. 1977. Volatile compounds from marine algae. Acc Chem Res 10:40–47. doi: 10.1021/ar50110a002. - DOI
-
- Kuhlisch C, Schleyer G, Shahaf N, Vincent F, Schatz D, Vardi A. 2020. Viral infection of algal blooms leaves a halogenated footprint on the dissolved organic matter in the ocean. bioRxiv https://www.biorxiv.org/content/10.1101/2020.09.08.287805v1. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
