

Protist diversity and community complexity in the rhizosphere of switchgrass are dynamic as plants develop
- PMID: 33910643
- PMCID: PMC8082632
- DOI: 10.1186/s40168-021-01042-9
Protist diversity and community complexity in the rhizosphere of switchgrass are dynamic as plants develop
Abstract
Background: Despite their widespread distribution and ecological importance, protists remain one of the least understood components of the soil and rhizosphere microbiome. Knowledge of the roles that protists play in stimulating organic matter decomposition and shaping microbiome dynamics continues to grow, but there remains a need to understand the extent to which biological and environmental factors mediate protist community assembly and dynamics. We hypothesize that protists communities are filtered by the influence of plants on their rhizosphere biological and physicochemical environment, resulting in patterns of protist diversity and composition that mirror previously observed diversity and successional dynamics in rhizosphere bacterial communities.
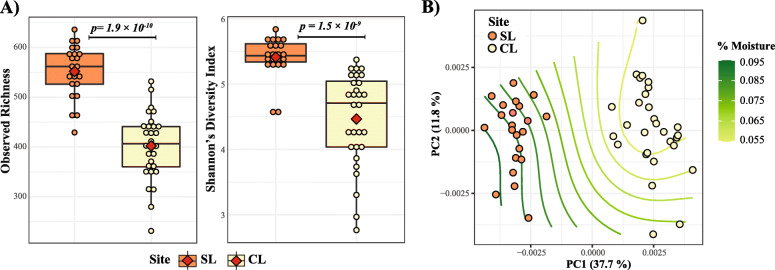
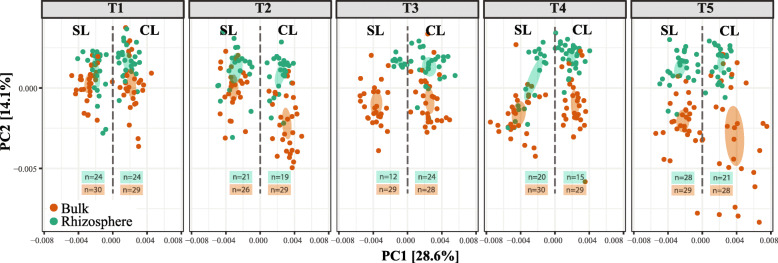
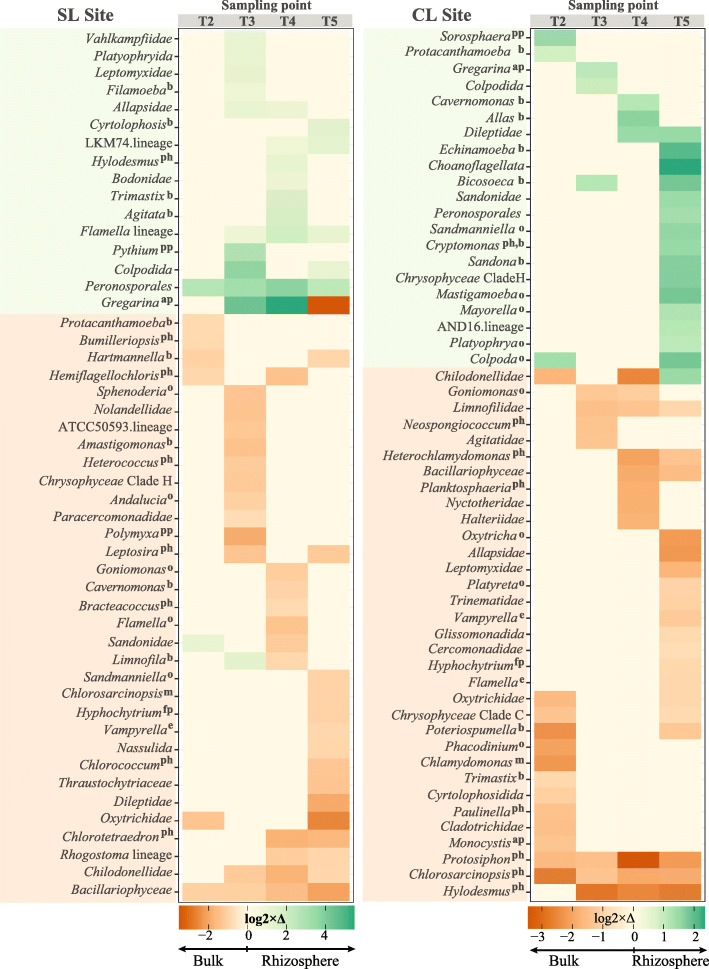
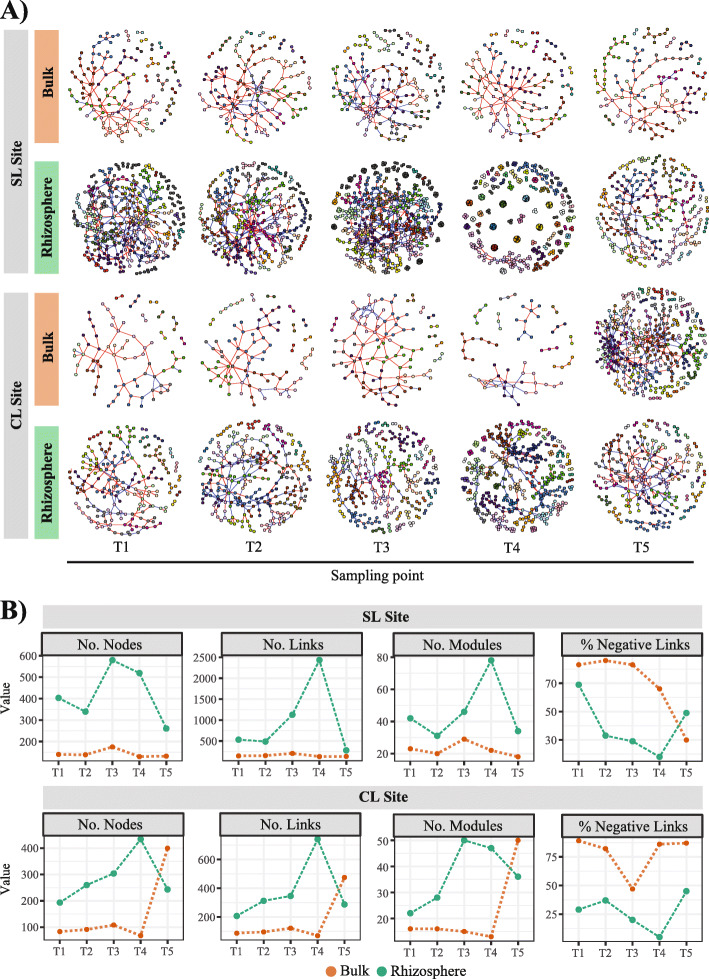
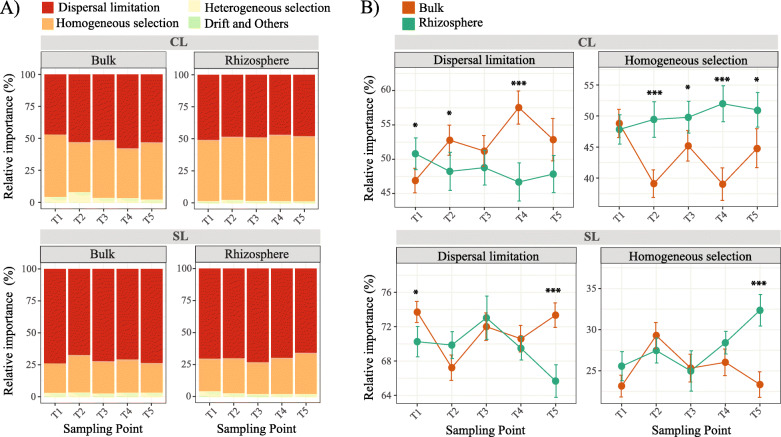
Results: We analyzed protist communities associated with the rhizosphere and bulk soil of switchgrass (SG) plants (Panicum virgatum) at different phenological stages, grown in two marginal soils as part of a large-scale field experiment. Our results reveal that the diversity of protists is lower in rhizosphere than bulk soils, and that temporal variations depend on soil properties but are less pronounced in rhizosphere soil. Patterns of significantly prevalent protists groups in the rhizosphere suggest that most protists play varied ecological roles across plant growth stages and that some plant pathogenic protists and protists with omnivorous diets reoccur over time in the rhizosphere. We found that protist co-occurrence network dynamics are more complex in the rhizosphere compared to bulk soil. A phylogenetic bin-based null model analysis showed that protists' community assembly in our study sites is mainly controlled by homogenous selection and dispersal limitation, with stronger selection in rhizosphere than bulk soil as SG grew and senesced.
Conclusions: We demonstrate that environmental filtering is a dominant determinant of overall protist community properties and that at the rhizosphere level, plant control on the physical and biological environment is a critical driver of protist community composition and dynamics. Since protists are key contributors to plant nutrient availability and bacterial community composition and abundance, mapping and understanding their patterns in rhizosphere soil is foundational to understanding the ecology of the root-microbe-soil system. Video Abstract.
Keywords: Community assembly; Rhizosphere; Soil microbiome; Soil protist; Switchgrass.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Heckman DS. Molecular evidence for the early colonization of land by fungi and plants. Science (80- ) 2001;293:1129–33. 10.1126/science.1061457 - PubMed
-
- Lambers H, Mougel C, Jaillard B, Hinsinger P. Plant-microbe-soil interactions in the rhizosphere: an evolutionary perspective. Plant Soil. 2009;321(1-2):83–115. doi: 10.1007/s11104-009-0042-x. - DOI
-
- Zhalnina K, Louie KB, Hao Z, Mansoori N, da Rocha UN, Shi S, et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat Microbiol. 2018;3(4):470–80. 10.1038/s41564-018-0129-3. - PubMed
