

Genome-Wide B Cell, CD4+, and CD8+ T Cell Epitopes That Are Highly Conserved between Human and Animal Coronaviruses, Identified from SARS-CoV-2 as Targets for Preemptive Pan-Coronavirus Vaccines
- PMID: 33911008
- PMCID: PMC8722450
- DOI: 10.4049/jimmunol.2001438
Genome-Wide B Cell, CD4+, and CD8+ T Cell Epitopes That Are Highly Conserved between Human and Animal Coronaviruses, Identified from SARS-CoV-2 as Targets for Preemptive Pan-Coronavirus Vaccines
Abstract
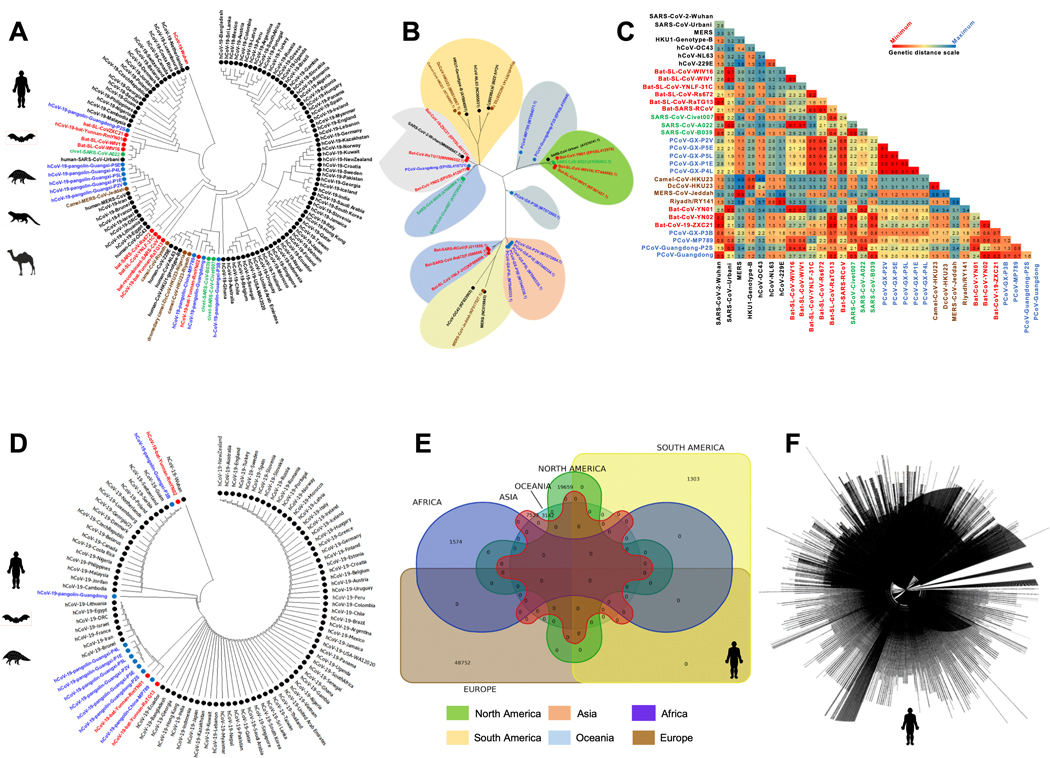
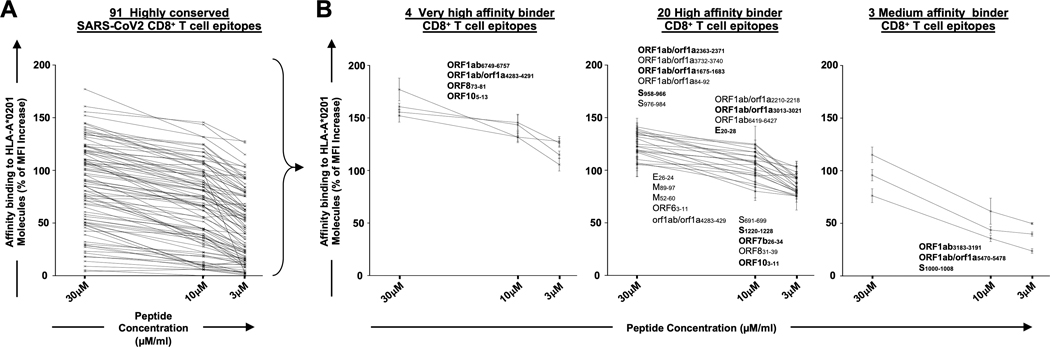
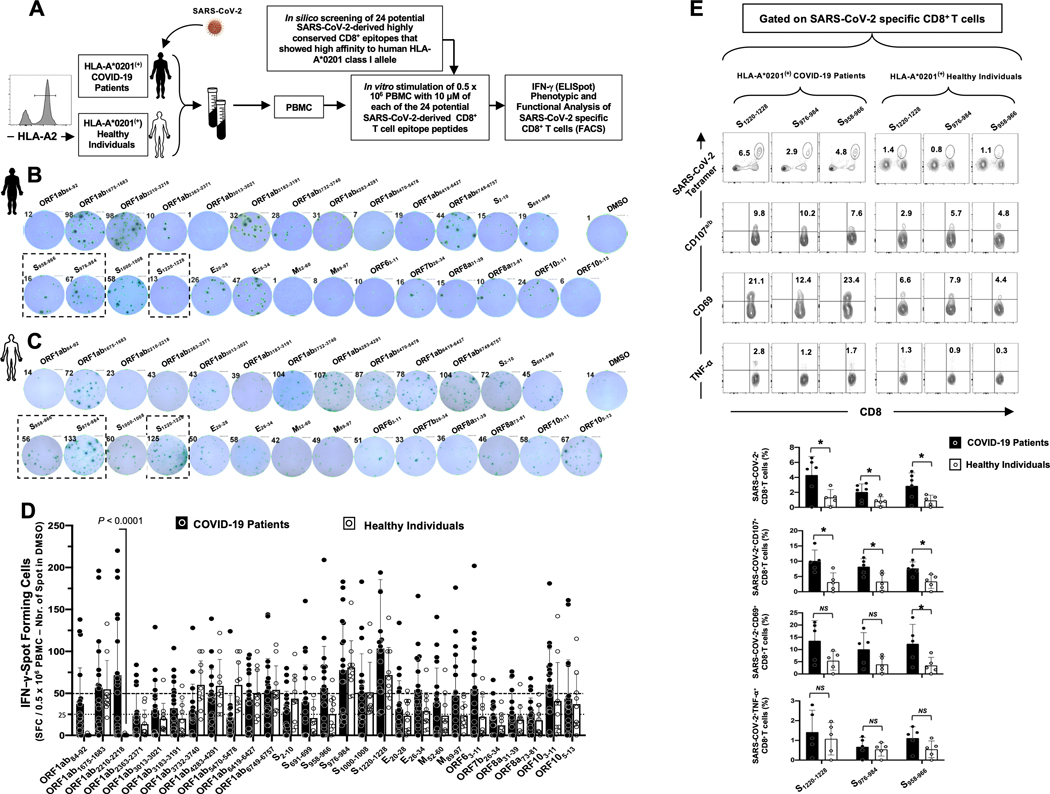
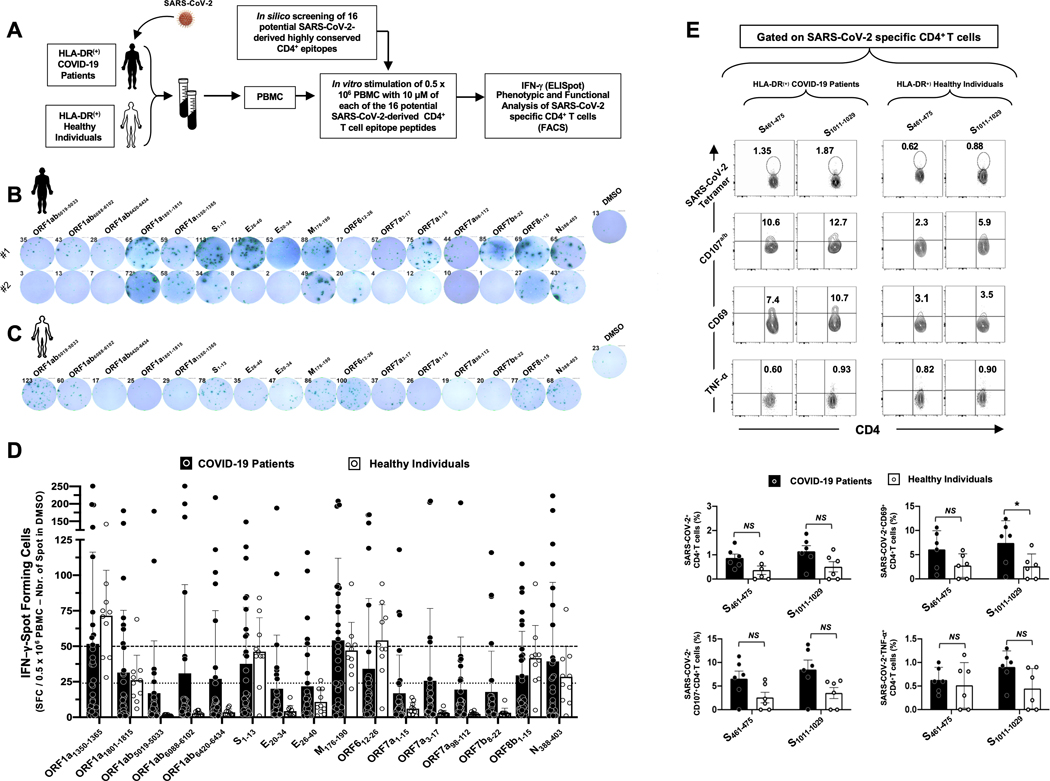
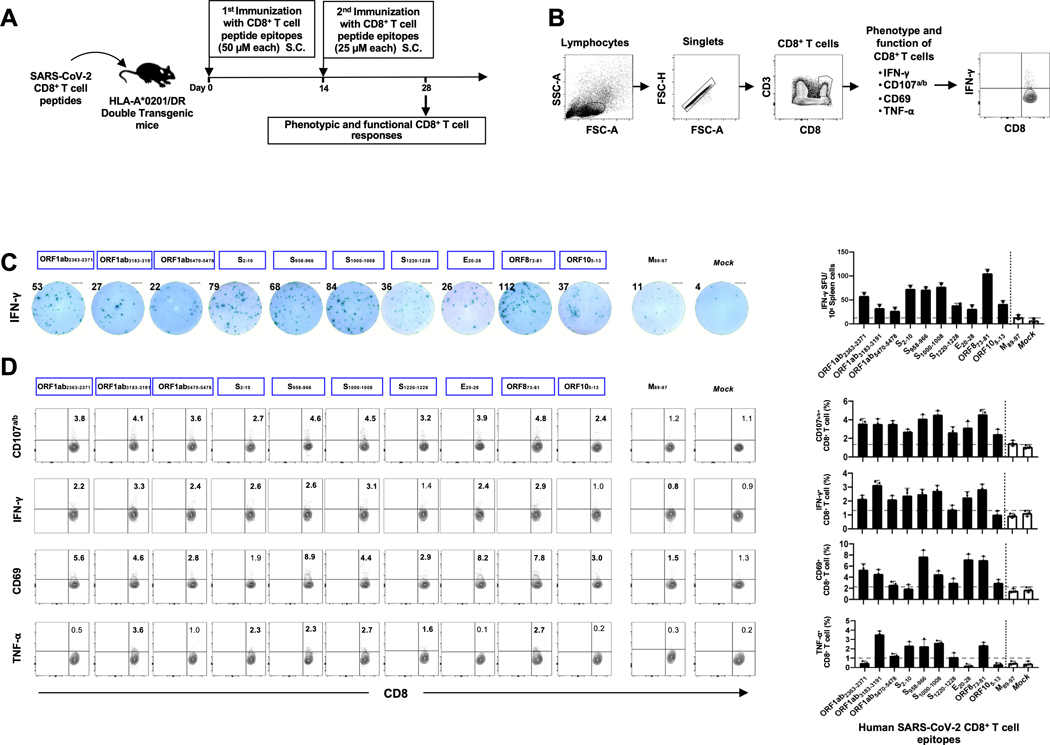
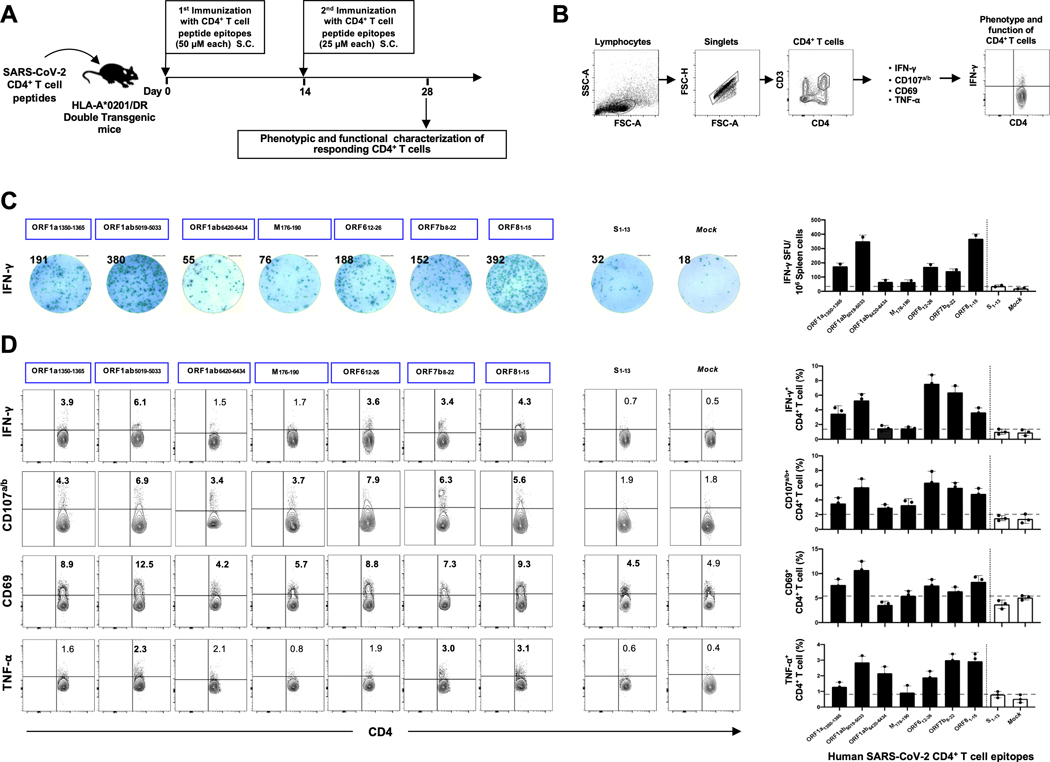
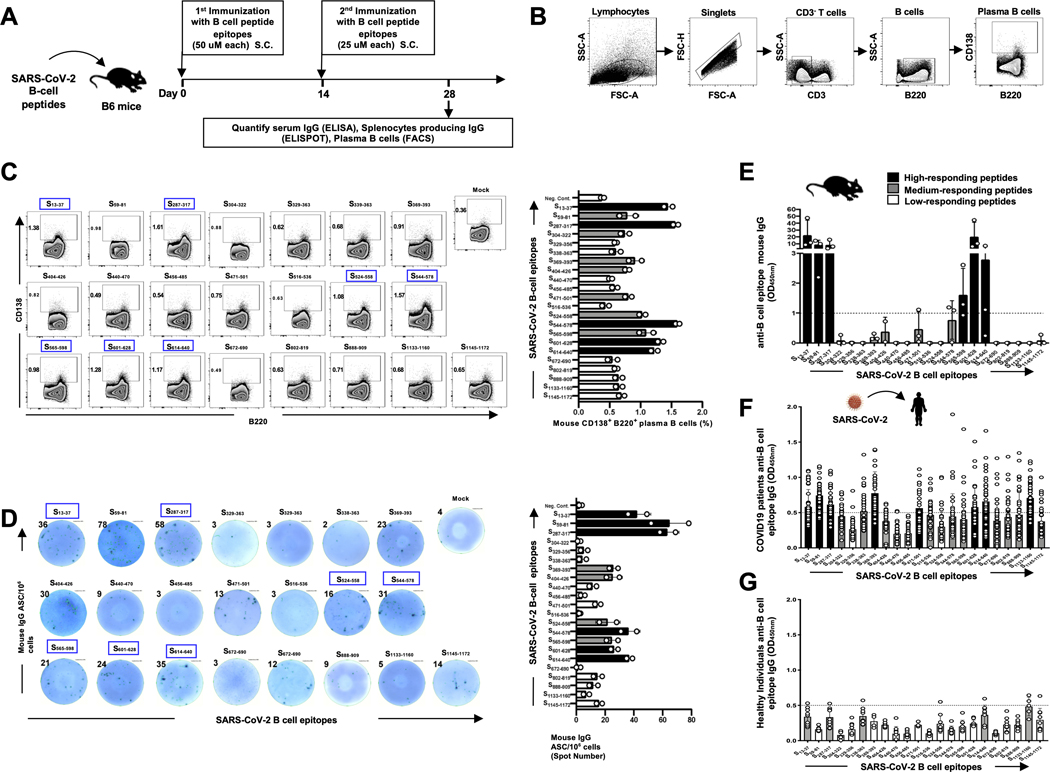
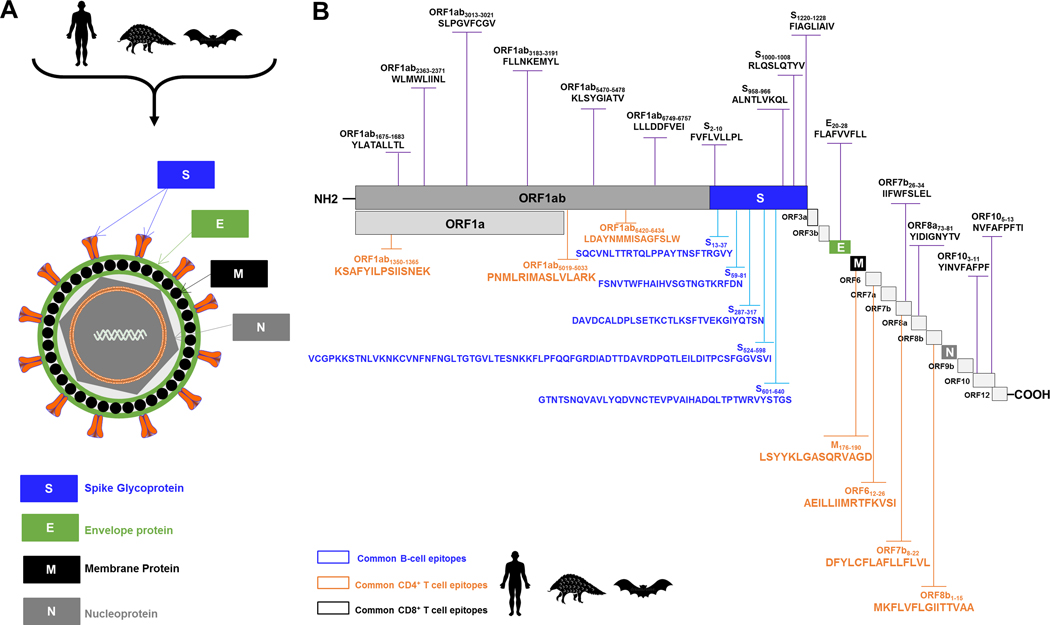
Over the last two decades, there have been three deadly human outbreaks of coronaviruses (CoVs) caused by SARS-CoV, MERS-CoV, and SARS-CoV-2, which has caused the current COVID-19 global pandemic. All three deadly CoVs originated from bats and transmitted to humans via various intermediate animal reservoirs. It remains highly possible that other global COVID pandemics will emerge in the coming years caused by yet another spillover of a bat-derived SARS-like coronavirus (SL-CoV) into humans. Determining the Ag and the human B cells, CD4+ and CD8+ T cell epitope landscapes that are conserved among human and animal coronaviruses should inform in the development of future pan-coronavirus vaccines. In the current study, using several immunoinformatics and sequence alignment approaches, we identified several human B cell and CD4+ and CD8+ T cell epitopes that are highly conserved in 1) greater than 81,000 SARS-CoV-2 genome sequences identified in 190 countries on six continents; 2) six circulating CoVs that caused previous human outbreaks of the common cold; 3) nine SL-CoVs isolated from bats; 4) nine SL-CoV isolated from pangolins; 5) three SL-CoVs isolated from civet cats; and 6) four MERS strains isolated from camels. Furthermore, the identified epitopes: 1) recalled B cells and CD4+ and CD8+ T cells from both COVID-19 patients and healthy individuals who were never exposed to SARS-CoV-2, and 2) induced strong B cell and T cell responses in humanized HLA-DR1/HLA-A*02:01 double-transgenic mice. The findings pave the way to develop a preemptive multiepitope pan-coronavirus vaccine to protect against past, current, and future outbreaks.
Copyright © 2021 by The American Association of Immunologists, Inc.
Figures
Update of
-
Genome-Wide Asymptomatic B-Cell, CD4 + and CD8 + T-Cell Epitopes, that are Highly Conserved Between Human and Animal Coronaviruses, Identified from SARS-CoV-2 as Immune Targets for Pre-Emptive Pan-Coronavirus Vaccines.bioRxiv [Preprint]. 2020 Sep 28:2020.09.27.316018. doi: 10.1101/2020.09.27.316018. bioRxiv. 2020. Update in: J Immunol. 2021 Jun 1;206(11):2566-2582. doi: 10.4049/jimmunol.2001438. PMID: 33024971 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
