

Subversion of Host Innate Immunity by Rickettsia australis via a Modified Autophagic Response in Macrophages
- PMID: 33912163
- PMCID: PMC8071864
- DOI: 10.3389/fimmu.2021.638469
Subversion of Host Innate Immunity by Rickettsia australis via a Modified Autophagic Response in Macrophages
Abstract
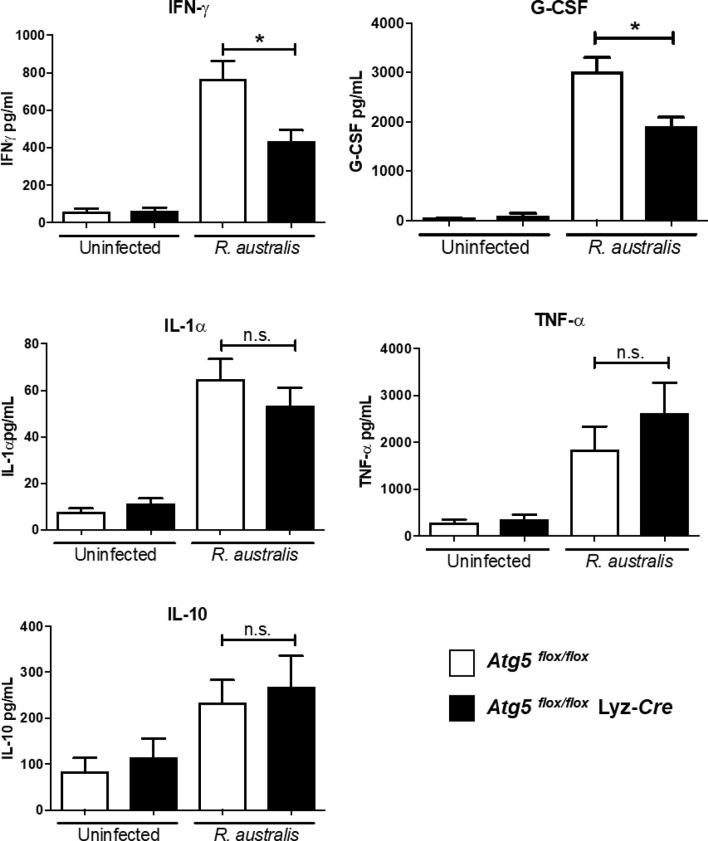
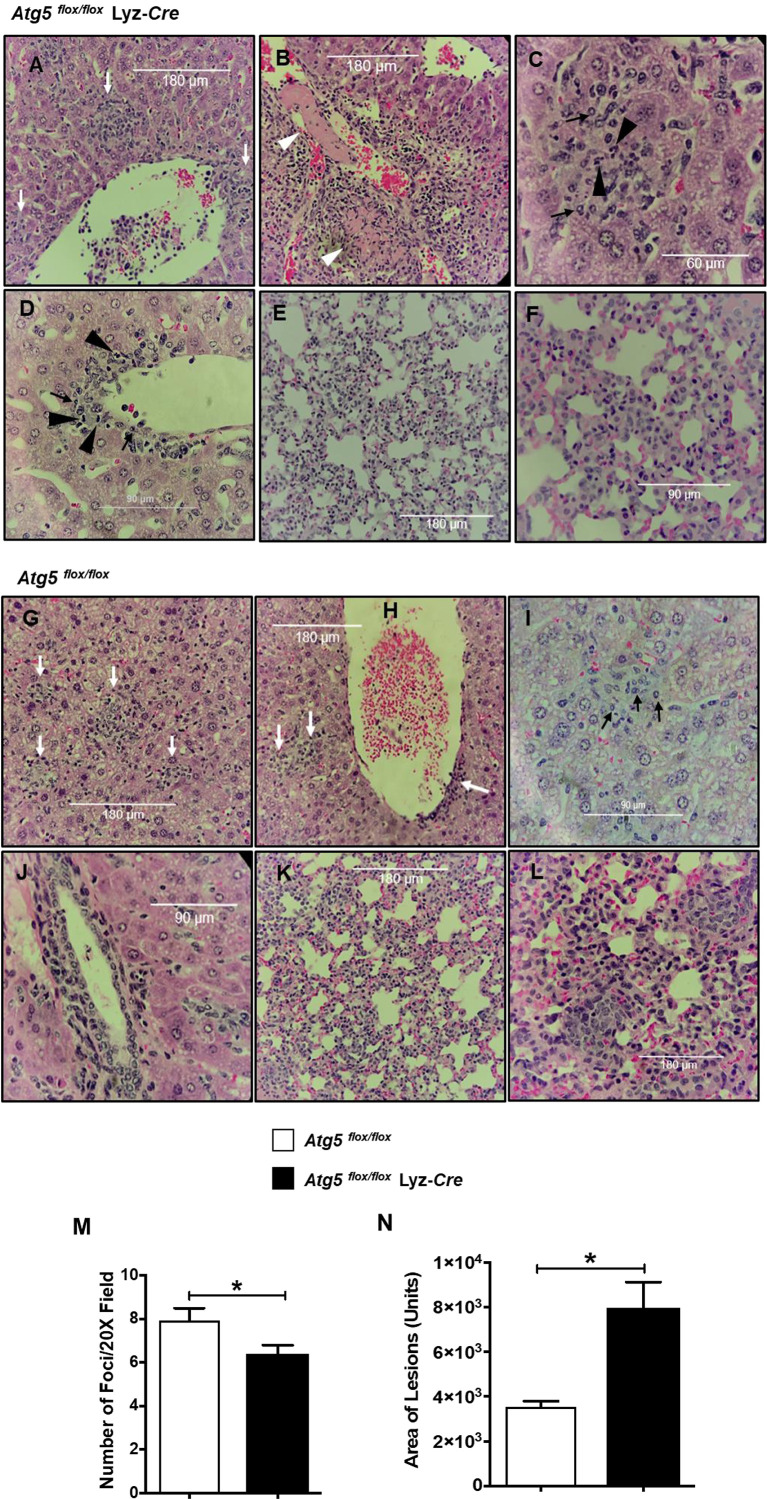
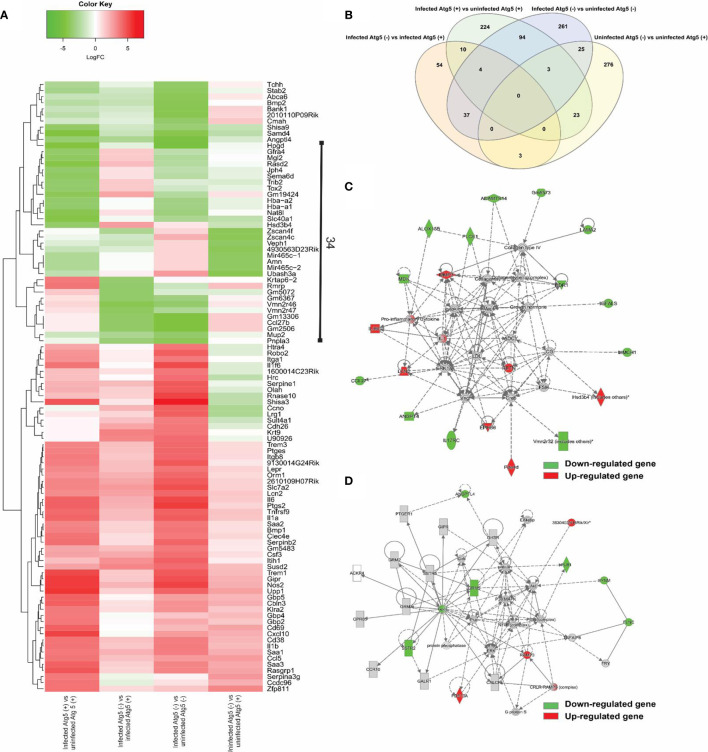
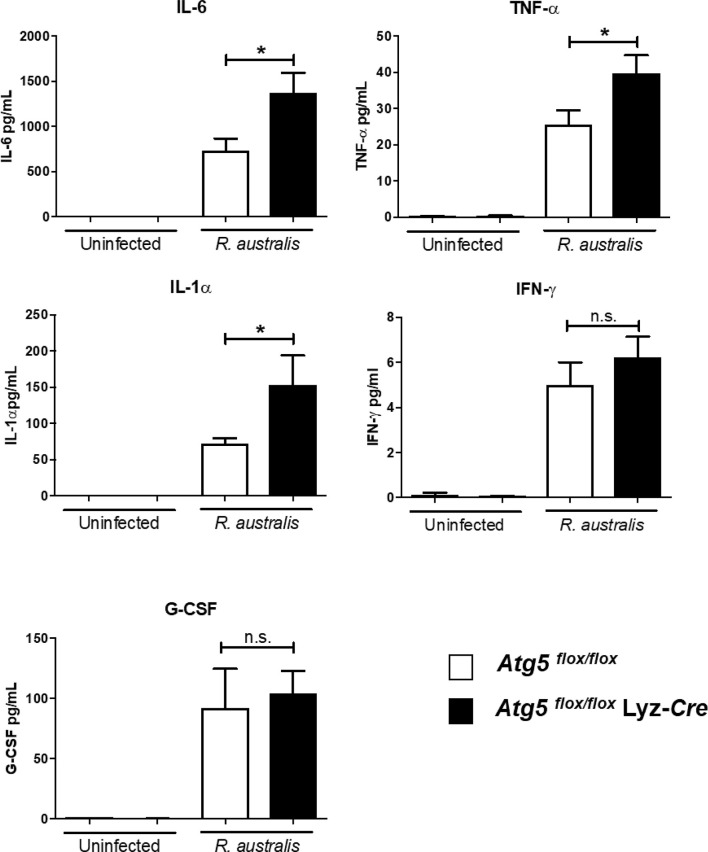
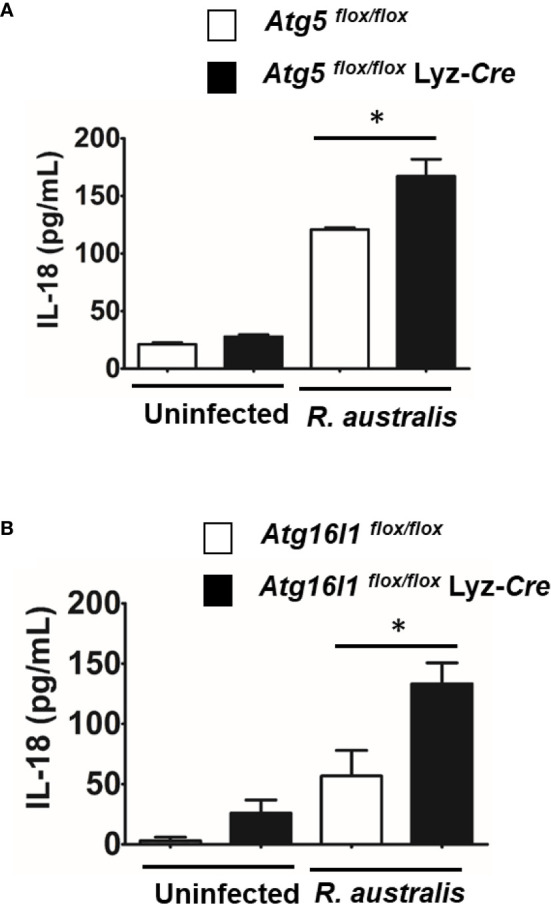
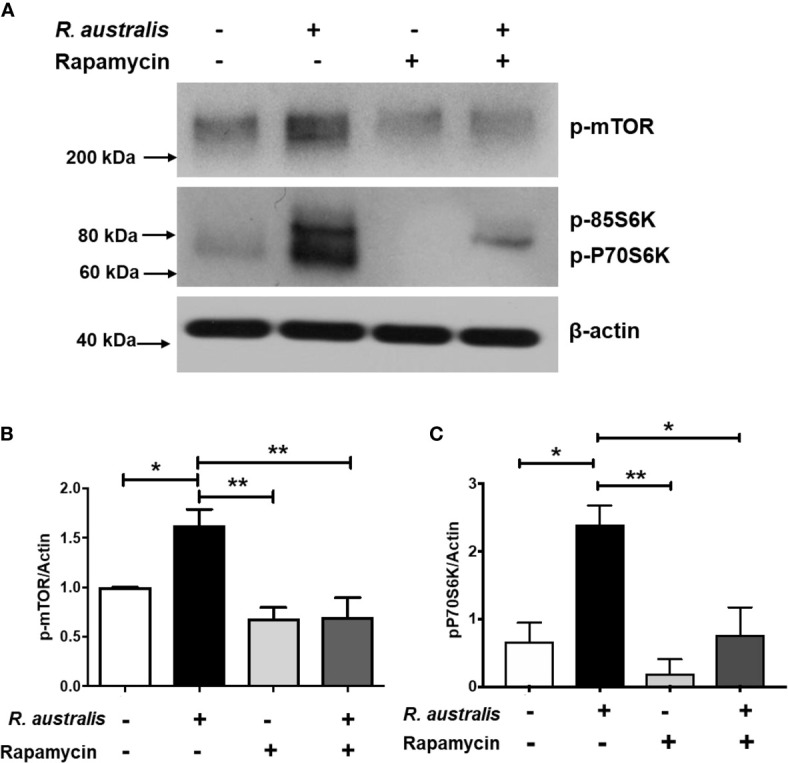
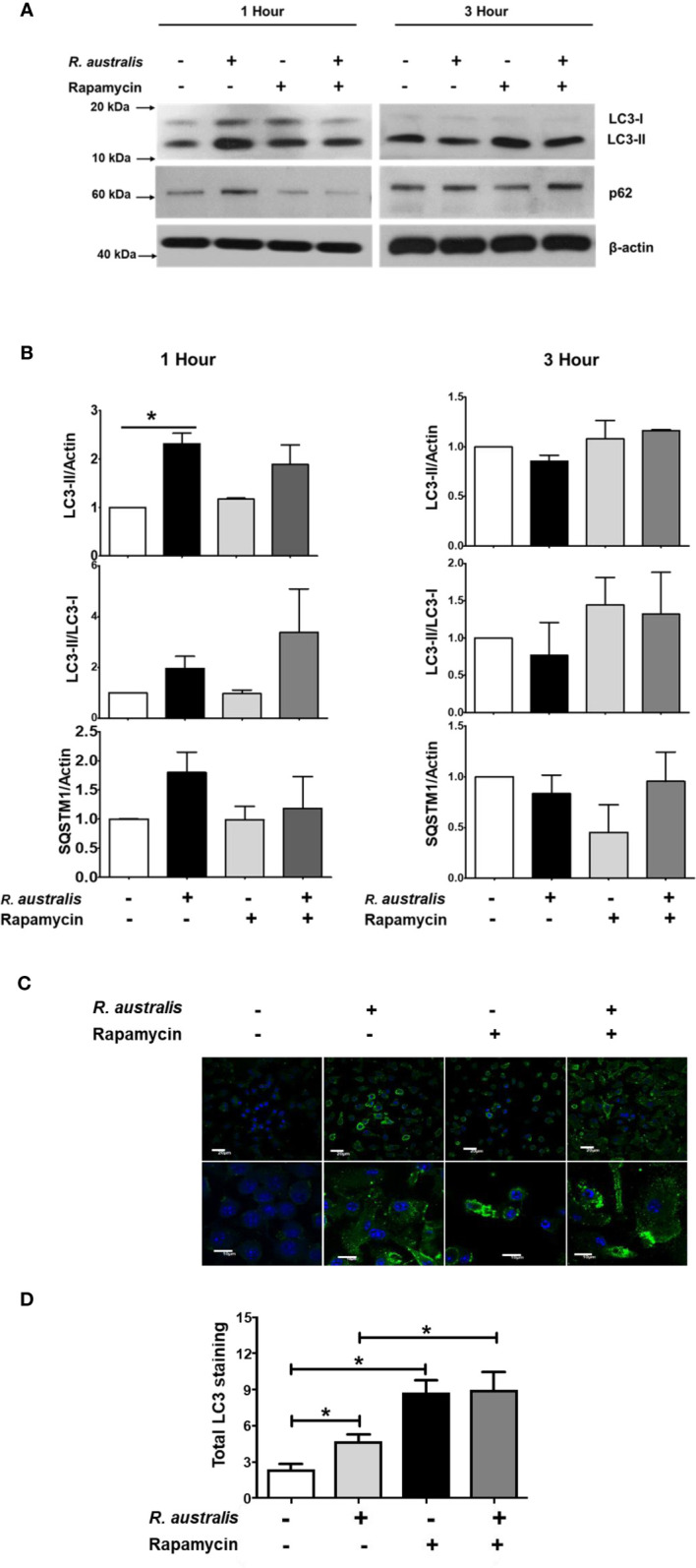
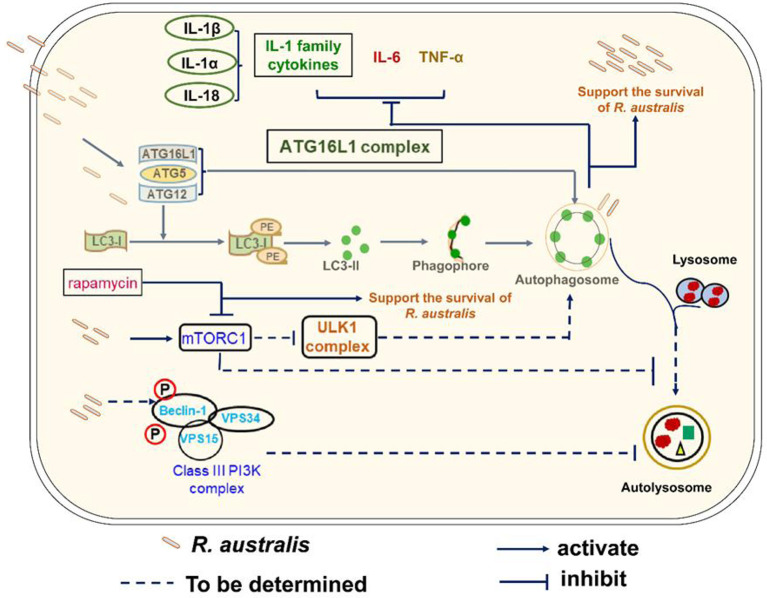
We recently reported that the in vitro and in vivo survivals of Rickettsia australis are Atg5-dependent, in association with an inhibited level of anti-rickettsial cytokine, IL-1β. In the present study, we sought to investigate how R. australis interacts with host innate immunity via an Atg5-dependent autophagic response. We found that the serum levels of IFN-γ and G-CSF in R. australis-infected Atg5flox/flox Lyz-Cre mice were significantly less compared to Atg5flox/flox mice, accompanied by significantly lower rickettsial loads in tissues with inflammatory cellular infiltrations including neutrophils. R. australis infection differentially regulated a significant number of genes in bone marrow-derived macrophages (BMMs) in an Atg5-depdent fashion as determined by RNA sequencing and Ingenuity Pathway Analysis, including genes in the molecular networks of IL-1 family cytokines and PI3K-Akt-mTOR. The secretion levels of inflammatory cytokines, such as IL-1α, IL-18, TNF-α, and IL-6, by R. australis-infected Atg5flox/flox Lyz-Cre BMMs were significantly greater compared to infected Atg5flox/flox BMMs. Interestingly, R. australis significantly increased the levels of phosphorylated mTOR and P70S6K at a time when the autophagic response is induced. Rapamycin treatment nearly abolished the phosphorylated mTOR and P70S6K but did not promote significant autophagic flux during R. australis infection. These results highlight that R. australis modulates an Atg5-dependent autophagic response, which is not sensitive to regulation by mTORC1 signaling in macrophages. Overall, we demonstrate that R. australis counteracts host innate immunity including IL-1β-dependent inflammatory response to support the bacterial survival via an mTORC1-resistant autophagic response in macrophages.
Keywords: Rickettsia; autophagy; innate immunity; mTOR signaling; macrophages.
Copyright © 2021 Bechelli, Rumfield, Walker, Widen, Khanipov and Fang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous