

Differential Responses of Cucurbita pepo to Podosphaera xanthii Reveal the Mechanism of Powdery Mildew Disease Resistance in Pumpkin
- PMID: 33912204
- PMCID: PMC8072357
- DOI: 10.3389/fpls.2021.633221
Differential Responses of Cucurbita pepo to Podosphaera xanthii Reveal the Mechanism of Powdery Mildew Disease Resistance in Pumpkin
Abstract
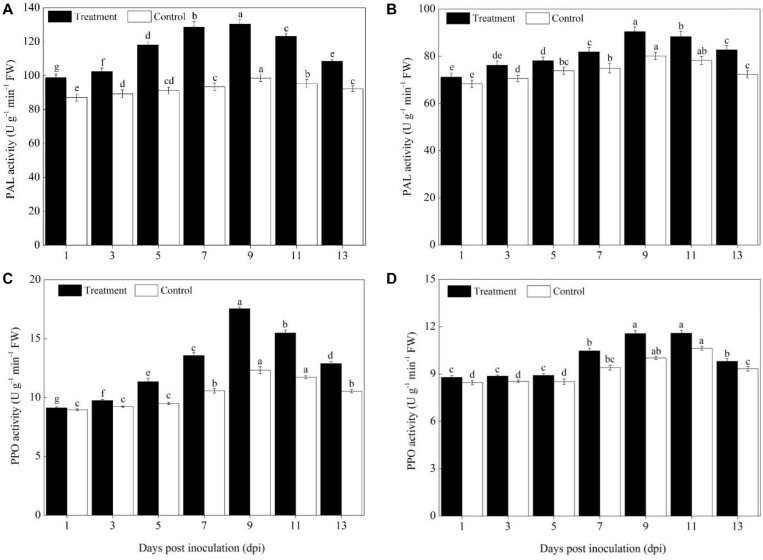
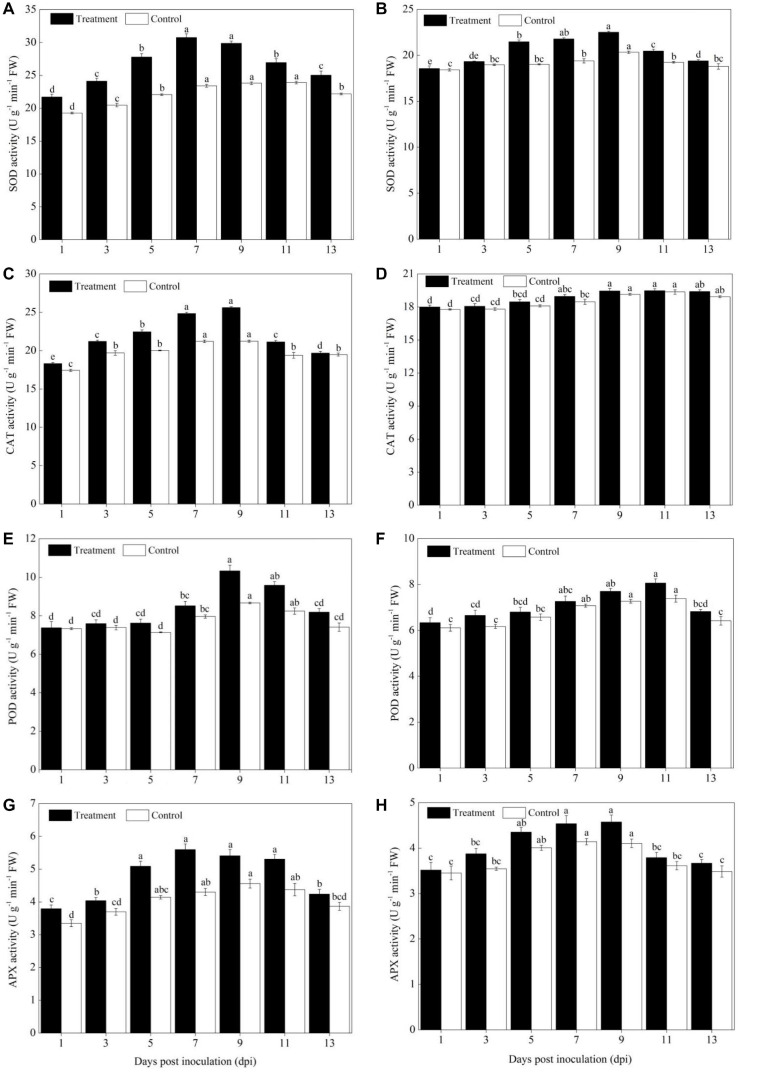
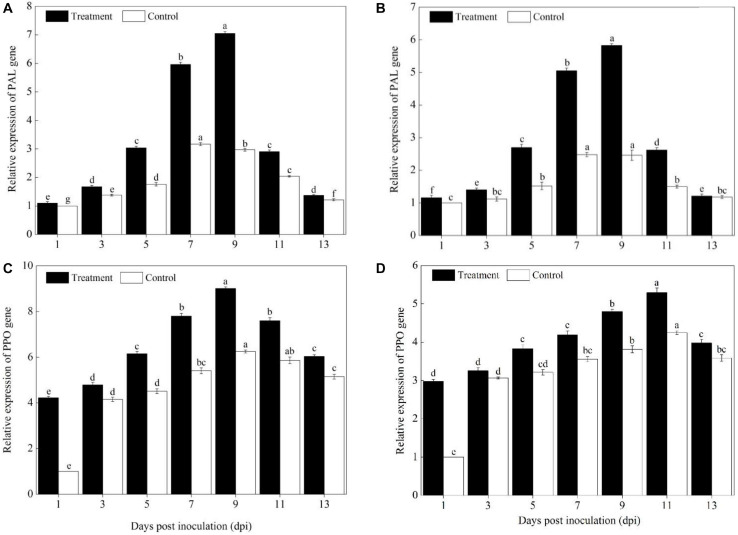
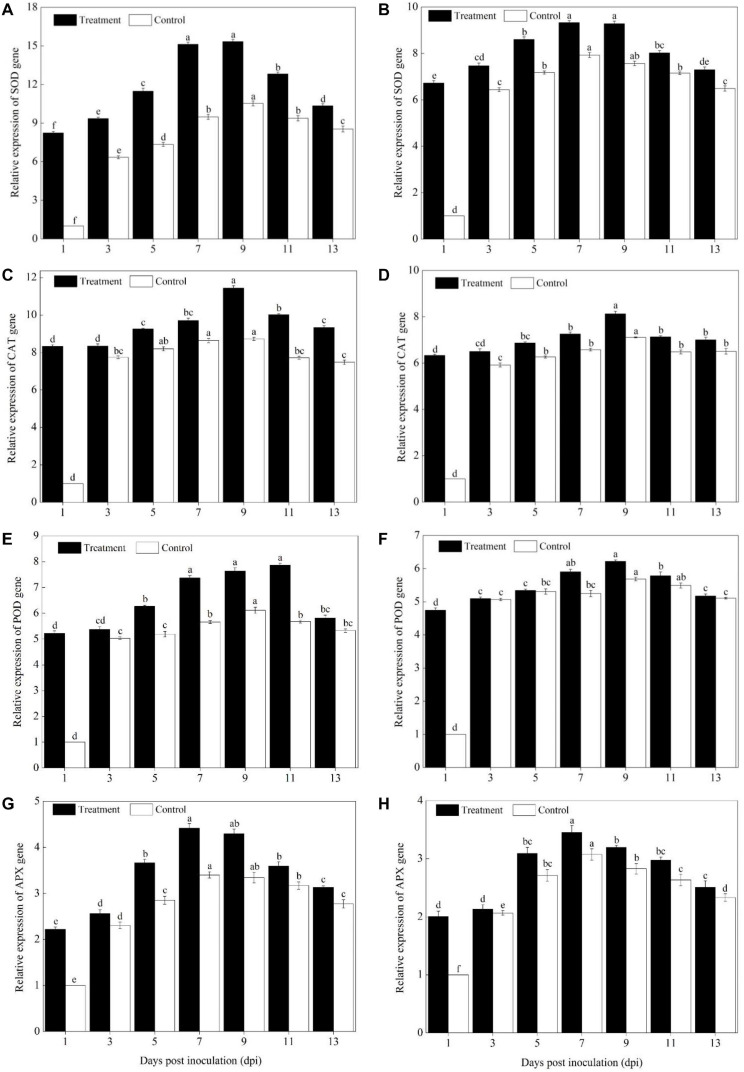
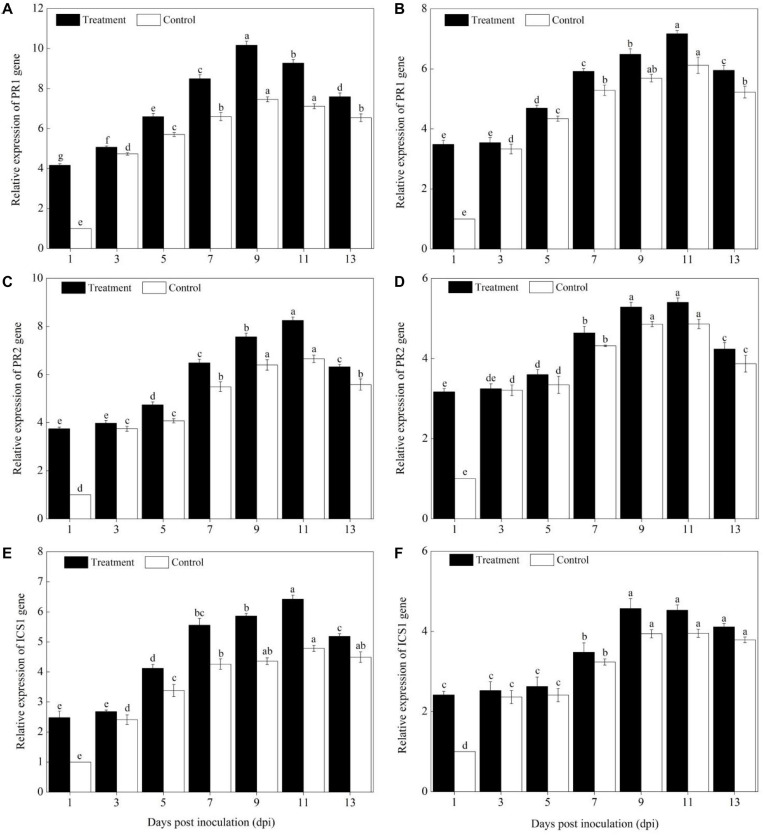
Powdery mildew is one of the most destructive diseases and the major cause to the production losses of cucurbit worldwide. A number of strategies have been developed and applied to discover suitable and safer methods to manage the occurrence of powdery mildew disease in pumpkins (Cucurbita pepo L.), but information is limited in screening tolerant germplasms and exploring their mechanisms in preventing the disease occurrence at physiological, biochemical, and molecular levels. Therefore, we investigated the responses of two commercial pumpkin cultivars to Podosphaera xanthii infection. Compared with mock-inoculated seedlings, few small and sparse powdery areas were observed on the leaves of the Sixing F1 cultivar on the 13th day after inoculation with P. xanthii, whereas a large number of diseased powdery areas and a layer of white powdery mildew were observed on the surface of Jin12 F1 leaves. The inoculation duration (7, 9, 11, and 13 days) significantly and continuously increased the disease incidence and index of pumpkin seedlings. The contents of H2O2, MDA, lignin, and total phenolics in the leaves of Sixing F1 and Jin12 F1 cultivars were markedly increased after inoculation with P. xanthii. However, the Sixing F1 cultivar exhibited much less reactive oxygen species (ROS) accumulation, a lower rate of lipid peroxidation, and a higher level of lignin and total phenolics contents after inoculation than the Jin12 F1 cultivar. Compared with untreated control pumpkin seedlings, significantly higher activities and gene expressions of the phenylpropanoids pathway enzymes (PAL and PPO), ROS scavenging defense enzymes (SOD, CAT, POD, and APX), and other salicylic acid (SA) signaling pathway marker genes were observed in the leaves of both cultivars after P. xanthii inoculation at different inoculation time points. These enhancements were significantly higher in Sixing F1 than Jin12 F1. Our results indicate that the Sixing F1 cultivar exhibited a much stronger ability in resistance to P. xanthii infection than the Jin12 F1 cultivar. Our results suggest that one possible mechanism of C. pepo cultivars to prevent the pathogen P. xanthii infection is by activating and enhancing the activity and gene expression of the phenylpropanoids pathway to synthesize phenolic substances and lignin, ROS scavenging defense enzymes to eliminate the harmful effects of ROS, and signaling pathway marker gene expression to improve plant disease resistance.
Keywords: Cucurbita pepo; Podosphaera xanthii; antioxidative defense system; disease incidence and index; gene expression; phenylpropanoids and SA pathways; powdery mildew; reactive oxygen species.
Copyright © 2021 Zhang, Liu, Xu and Zhou.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
-
- Abdel-Monaim M., Abdel-Gaid M., Armanious H. (2012). Effect of chemical inducers on root rot and wilt diseases, yield and quality of tomato. Int. J. Agric. Sci. 7 211–220.
-
- Babosha A. V., Ryabchenko A. S., Avetisyan G. A., Avetisyan T. V. (2020). Visualization of the halo region in plant-powdery mildew interactions by cryoscanning electron microscopy. J. Plant Path. 102 103–111. 10.1007/s42161-019-00387-3 - DOI
-
- Babu A. N., Jogaiah S., Ito S., Nagaraj A. K. (2015). Improvement of growth, fruit weight and early blight disease protection of tomato plants by rhizosphere bacteria is correlated with their beneficial traits and induced biosynthesis of antioxidant peroxidase and polyphenol oxidase. Plant Sci. 231 62–73. 10.1016/j.plantsci.2014.11.006 - DOI - PubMed
-
- Barickman T. C., Horgan T. E., Wilson J. C. (2017). Efficacy of fungicide applications and powdery mildew resistance in three pumpkin cultivars. Crop Prot. 101 90–94. 10.1016/j.cropro.2017.07.025 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous